HONR 259C "Fearfully Great Lizards": Topics in Dinosaur Research
Fall Semester 2019 "Were Dinosaurs Warm-Blooded?" (Dinosaur Physiology)
The fuzzy neornithischian Kulindadromeus by Andrey Atuchin (2014)
Key Points:
•Modern animals are often characterized as "warm-blooded" (mammals, birds) and "cold-blooded" (everything else)
•This is a simplification of several related phenomena: energy source (endothermy vs. ectothermy); metabolic rate (tachymetabolism vs. bradymetabolism); and temperature stability (homeothermy vs. poikilothermy)
•Dinosaur species were initially inferred to be "cold-blooded", but similarity in posture and other traits led Owen to suggest they might have been warm-blooded; since that time various researchers have examined the alternatives.
•Dinosaur posture, locomotion, feeding adaptations, growth rates, bone texture, inferred respiration, and predator-prey ratios point to elevated metabolisms relative to today's non-avian sauropsids.
The Hot-Blooded Dinosaurs: Reconstructing Dinosaur Physiology
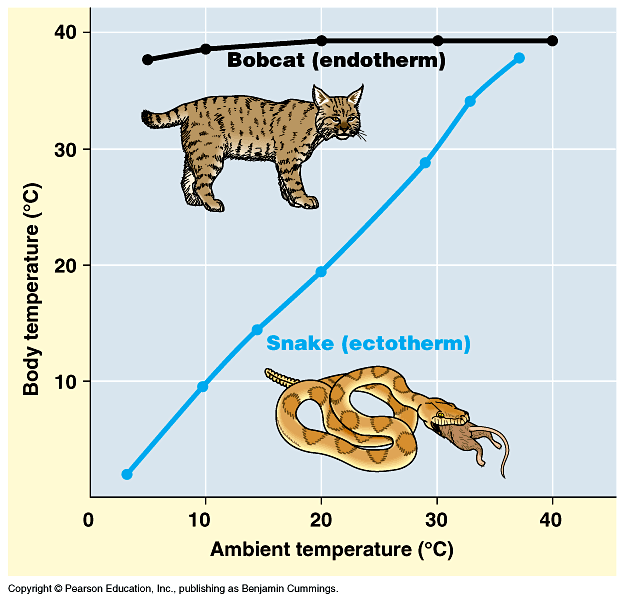
Among modern vertebrates, some gross generalizations: Birds and
mammals are warm-blooded; that is, they are warmer than the environment around them in typical temperate and colder environments. Crocodilians, lepidosaurs, turtles, amphibians, most fish, and almost all invertebrates are cold-blooded: their bodies are generally only about as warm as the general environment around them, so consequently they feel cool to the touch outside of tropical situations; in contrast, warm-blooded animals have temperatures largely independent of the outside temperature,
so they feel warm to the touch. Need to be precise as to definitions of terms. "Warm-blooded" and "Cold-blooded" actually encompass several different (although related) topics:
Energy Source: whence comes the majority of the energy to "run" the animal?
In "Warm-blooded" animals, the main energy source are extra mitochondria whose main purpose is to convert food energy to heat energy: called endotherms ("inside heat")
Metabolic Rate: how much food energy ("fuel") is used up over time?
In "Cold-blooded" animals, rate of fuel usage is low: called bradymetabolic ("slow metabolism")
In "Warm-blooded" animals, rate of fuel usage is HIGH: called tachymetabolic ("fast metabolism")
Temperature Variation over Time: how stable is the body temperature over time?
In "Cold-blooded" animals, body temperature fluctuates with the external environment: called poikilotherms ("fluctuating heat")
In "Warm-blooded" animals, body temperature regulated by internal mechanisms and thus more stable: called homeotherms ("same heat")
A typical cold-blooded animal is an ectothermic bradymetabolic poikilotherm: needs to get its energy from the sun and fluctuates with external environment (but can moderate fluctuations by moving from sunlight to shade and vice versa); however, needs very little
food (snakes can go weeks without feeding, for example). Cold blooded animals become torpid at night and in colder weather.
A typical warm-blooded animal is an endothermic tachymetabolic homeotherm: its body temperature is stable and activity levels can remain high for long periods of time, at night, and in colder weather; however, needs a LOT of food or will die (imagine the
effects of not feeding a cat or dog for weeks!).
Additional issues to consider:
Resting metabolic rate (RMR) vs. active metabolic rate (AMR): "warm-blooded animals tend to have RMRs 4-10x that of similar sized "cold-bloods", but AMR is similar in both
Duration of sustained activity: "warm-blooded" animals tend to have longer durations of sustained activity
Recovery time between periods of activity: often much shorter for "warm-blooded" animals
Why evolve such an expensive trait as endothermy? Some suggestions have included:
Increased aerobic capacity, allowing for greater total activity levels and greater ability to recover from sustained activity
Greater environmental tolerance: endotherms can live in wider range of latitudes and altitudes
Increased metabolic efficiency due to homeothermy: can "fine-tune" physiological systems to a narrow range of temperatures
Increased ability for parental care: both brooding/gestating at constant temperature, and increased ability to watch over young
Note that it is not just mammals and birds that are "warm-blooded". For example, tunas, billfish (sailfish, swordfish, marlins), lamniform sharks (like great whites and makos), boid snakes (pythons, etc.; but only while brooding), and certain plants (which aren't "blooded" as such, but some can emit internally-generated heat).
When dinosaurs were first discovered, they were interpreted as being no more than gigantic cold-blooded lizards. However, as early as 1842 Owen (in the very paper in which he named "Dinosauria") speculated that dinosaurs may have been warm-blooded like mammals. During most of the 20th Century the model of dinosaurs as cold-blooded returned. Work by John Ostrom (of Yale University) and his colleagues and students (especially Robert Bakker) presented new information that dinosaurs were in fact warm-blooded. This hypothesis generated considerable research (both in support and in attempts to falsify it): this change in thinking about dinosaurs and renewed interest in dinosaurian studies has been termed the "Dinosaur Renaissance".
Among the lines of evidence supporting dinosaurian warm-bloodedness:
Bone histology (microscopic analysis of tissue and cellular structure) can test between bradymetabolic and tachymetabolic organisms. Bone becomes reworked (that is, removed then redeposited) as a normal part of vertebrate physiology, as bone is a major store for nutrients like calcium and phosphorus. Bradymetabolic animals show little sign of reworking, as their slower metabolism does not need as much nutrients as quickly. In contrast, tachymetabolic animals show considerable reworking. Examination of the bones of dinosaurs show a high degree of reworking, even as juveniles.
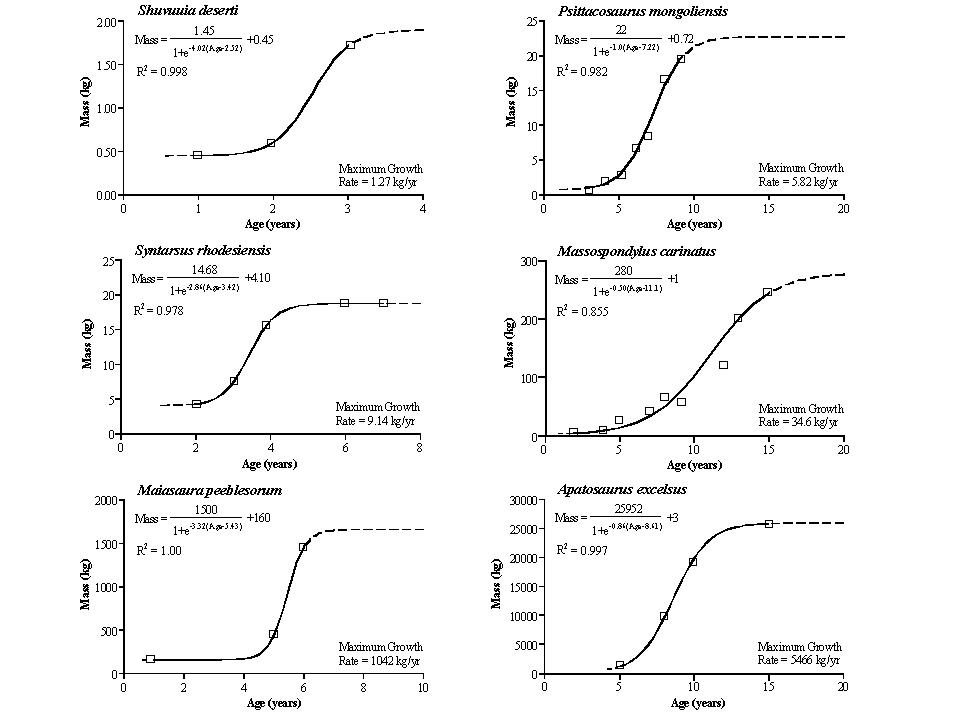
Using skeletochronology, the maximum rate of growth of dinosaurs can be calculated. When plotted against body size, it is found that dinosaurs had much higher growth rates than ectotherms of the same size; in fact, the dinosaur growth rate is about the same as mammals and ground-dwelling birds.
Texture of bone also shows signs of the rate of growth. Dinosaurs show fast rates similar to mammals; pseudosuchians and other archosauriforms show an intermediate rate; and ectotherms show a slow growth rate.
Computer models of the required metabolic rates required for even walking and slow running for large bipedal dinosaurs exceeds the metabolic rate of ecotherms. So if they actually moved, big theropods HAD to have been endotherms.
And the data for small dinosaurs and dinosauromorphs are at least consistent with endothermy for walking, and require endothermy if they were even moderate runners (as their skeletons and foot print evidence shows).
Comparison of the oxygen isotope data of dinosaurs with known ectotherms of the same environment (modern-style crocodilians, turtles, and amphibians) finds that the dinosaurs show lower body temperatures in the hottest environments and lower body temperatures in moderate and colder environments. This suggest that dinosaur body temperatures were independent of the local environment.
Predator-Prey ratios (first considered by R.T. Bakker looked at the trophic relationships in communities to try and determine the thermophysiology of dinosaurs and other extinct forms. His technique:
In modern endothermic communities very few predators compared to many herbivores (tachymetabolic predators require a lot of food, so only a few can survive in a given region).
Bradymetabolic predators require a lot less food, so same amount of potential food can support many bradymetabolic predators.
In order to calculate P/P ratios, Bakker had to consider the different sizes of the various populations. Used biomass (# kgs or tons of flesh) rather than number of individuals
Found that modern populations had P/P ratios of 0.5-4 %
Looking at fossil record, found:
Basal synapsid-dominated faunas of the Early Permian: 25-30%, much higher than modern populations. Most paleontologists have accepted this as a cold-blooded community
Therapsid-dominated faunas of the Middle and Late Permian and earliest Triassic: 10-20%, seemingly between endo- and ectothermic
populations
Pseudosuchian-dominated faunas of the Middle and Late Triassic: 10-20%, as in therapsid communities
Dinosaur-dominated faunas of the Jurassic and Cretaceous: 0.5-3.5%, as in modern endotherms!
Mammal-dominated faunas of the Cenozoic: 0.5-4.5%, known endotherms
Let's consider the equations of life. First, the aerobic respiration equation, the primary means by which animal cells operate:
C6H12O6 + 6O2 yields 6CO2 + 6H2O + Energy
(That is, food (glucose) plus oxygen yields waste carbon dioxide and waste water, plus energy).
If an animal's cells can't get enough oxygen, there is a second way of getting energy: the anaerobic respiration equation:
C6H12O6 yields 2C3H6O3 + Energy
(That is, food yields lactic acid plus energy (although much less than the aerobic respiration.) Lactic acid itself needs oxygen to break down, so you cannot run on anaerobic respiration for very long.
If you want to evolve endothermy, you need to:
Increase glucose intake, plus...
Increase oxygen intake, plus...
Increase the speed of distribution of glucose and oxygen throughout the body, plus...
Deal with excess carbon dioxide, water, and heat.
So, where do we stand on dinosaur metabolism?
All living dinosaurs (Aves) are endothermic tachymetabolic homeotherms
The living outgroups (crocodilians, lepidosaurs, turtles) are all ectothermic bradymetabolic heterotherms
Non-avian dinosaurs show many anatomical features suggesting levels of activity higher and/or more continuous than that seen in modern "cold-blooded" animals
Non-avian dinosaurs show growth patterns comparable to those of modern endotherms, and unlike those of modern and extinct ectotherms
What would be necessary to justify the above observations?
Non-avian dinosaurs would need active ventilation (breathing) to power the muscles and to fuel the growing tissue
Non-avian dinosaurs would need strong, active heart to get the oxygen to the muscles and tissues
Non-avian dinosaurs would need structures to control heat
Is there evidence for these features in dinosaurs? YES!:
Dinosaur Respiration:
Primitive tetrapods are subject to Carrier's Constraint: the same muscles for breathing are used for locomotion (bending from side-to-side). Thus, most primitive tetrapods tend to hold their breath while breathing, meaning that they rely more on anaerobic respiration and require long
recovery intervals.
Mammal-style diaphragm breathing is an advanced therapsid feature; most tetrapods breath by gulping air and by rib breathing
Specialized lizards developed neck breathing separate from rib breathing
Living dinosaurs (birds) have extremely specialized breathing:
Pump their lungs by rocking their hips up and down
Crocodilians have their own specialized breathing:
Pubis is mobile, and rocks back and forth pushing & pulling the liver
Functions like the mammalian diaphragm, to have additional active breathing
Recent analysis shows that they, like birds, had unidirectional flow through their lungs (with passages rather than airsacs)
Speculation: belly breathing is an archosaurian synapomorphy:
In primitive archosaurs, primitive pseudosuchians, and most dinosaurs other than birds, muscles from the pelvis would pull gastralia down, which would inflate the lungs
This would give these animals extra oxygen for their metabolism
Becomes modified in crocodilians (liver pump with mobile pubis), pterosaurs (a mobile "prepubis"; another liver pump?), birds, and ornithischians (mobile pubes or other parts of the pelvis in some ornithischian groups)
Additionally, the air flow in all archosaurs would have been unidirectional
Furthermore, strong evidence that theropods and sauropods (at least) had air sacs like those of birds:
Chambers in vertebrae are very similar to those of birds
Air sacs may have been present in other dinosaurs, but apparently did not enter the vertebrae
Dinosaur Hearts:
Turtles and lepidosaurs have three chambered hearts
Birds and mammals have four chambered hearts:
A "double pump" system, so the heart acts as a control between lungs and body
Shunts blood to lungs before going out to body, so all the blood getting to the tissues are fully oxygenated
Also, can allow these animals to be taller, since the heart pressure control separates lungs and body, and therefore pressure on lung blood vessels won't get too high
Crocodilians actually have specialized (NOT primitive) four-chambered hearts:
Operate as four-chambered heart on land, shifts to two chambered underwater since doesn't need to get blood to lungs
Since both birds and crocodilians have four-chambered hearts, assumption is that all extinct archosaurs, including non-avian dinosaurs, did too
What about controlling heat?
Keeping the Heat In: Insulation Issues: One problem that small-bodied organisms encounter is the fact that a small organism has a much higher surface area/volume ratio than a large one. Because of this, small animals tend to lose heat much faster than big ones. In contrast, large animals lose heat to or gain heat from the outside world only gradually. This has led some people to suggest the possibility that large dinosaurs exhibited "gigantothermy": effective homeothermy achieved because of large body size. However, this would not apply to small-bodied dinosaurs: either adults of small species or the hatchlings of giants. So how could these keep warm?
There is strong evidence that many--if not most--of the theropods had a fuzzy body insulation over the body: true feathers in the advanced groups, simpler "protofeathers" in the primitive ones. Such fuzz would help keep the warmth in the body. In fact, this is the primary function of the fur of mammals, and one of the functions of body feathers in birds. The recent discovery of 1.4 t Yutyrannus demonstrates that even some giant theropods were fuzzy.
Recent discovery of the early Late Jurassic Chinese ornithischian Tianyulong and the similar aged Kulindadromeus of Siberia showed they too had a fuzzy body covering over at least part of its body! If this is found to be homologous to the protofeathers of tetanurine theropod saurischians it would suggest that the concestor of all dinosaurs was fuzzy, and that dinosaurs were thus fuzzy ancestrally! (In the case of Kulindadromeus, there are also also scales, plates, and additional bizarre tufted plates.) At present, however, there is enough uncertainty to make the homology between Tianyulong's fuzz, Kulindadromeus's diverse integument, and theropod protofeathers suspicious. (But do not be terribly surprised if in the future we discover that most dinosaurs were fuzzy to some degree or another! All we need is a fuzzy primitive sauropodomorph, and it is basically a done deal!)
Dumping Heat: Mechanisms to Remove Heat:
Some dinosaurs have conspicuous large sails or plates or frills or long necks or long tails that might have been used to dump waste heat
However, other structures may have also been used to regulate temperature:
The antorbital fenestra (also the promaxillary and maxillary fenestrae of various theropods) housed soft tissue air sacs
These air sacs may have been useful to transport waste heat
Also, many larger dinosaurs have enlarged and/or elaborate nares
These may have been useful in dumping waste heat
The enlarged narial regions may support tissues for a different function: recovery of moisture. In living endotherms, rapid rate of respiration would dry out lungs if not for some specialized tissues called nasal turbinates:
Rare or small in modern ectotherms
Extremely large in mammals, where they support thin tissues which trap moisture going out, and rewets on way back in
Fairly large in many birds, but are cartilage rather than bony
Also, some birds seem to rely on air sac system for this purpose
Most non-avian dinosaurs do not show much evidence for internal nasal turbinates, but the air sac system and/or tissues in the enlarged narial regions of bigger dinosaurs may have served this function
An secondary advantage of using respiratory turbinates to dump heat is that it helps direct some of the air flow onto the part of the nasal chamber associated with olfaction:
It has just recently been recognized that the expanded frontopareital fossa (anteriorly-expanded sections of the supratemporal fenestrae) were filled by vascular tissue and fat rather than by muscle. So these areas may have helped to radiate heat to keep the brains of dinosaurs cool (they seem to have that function in modern crocodilians):
Some Complications
Is this whole debate a false dichotomy?
Is everything EITHER an endothermic tachymetabolic homeotherm OR an ectothermic bradymetabolic poikilotherm?
Some additional possibilities:
Gigantothermy:
From geometry, as linear dimensions double, the surface area goes up by squares, and the volume by cubes:
Side Length
Surface Area (SA)
Volume (V)
SA/V
1
6
1
6/1 = 6
2
24
8
24/8 = 3
3
54
27
54/27 = 2
4
96
64
96/64 = 1.5
As size increases, SA/V decreases.
The mass of an animal, and the heat it produces, is based on its volume.
The rate at which an animal can gain and lose heat is based on its surface area.
Therefore, with bigger body size it takes longer and longer for heat to be lost or gained:
Become homeothermic without having the energy costs of endothermy!
Problem: No good living models (elephants once thought to be partial gigantotherms, but does not now seem true; marine leatherback turtles are, but are not in same type of environment)
Also, gigantothermy might apply to large dinosaurs, but would not apply to small species or to babies.
Heterometabolism:
Changeable metabolic rate: tachy- to bradymetabolic.
Two main types: behavioral and ontogenetic:
Behavioral heterometabolism:
Normally operate as bradymetabolic, but shift into "high gear" in certain circumstances
Living examples: sharks in feeding frenzy; pythons while brooding
Some have suggested that specialized breathing structures may have let dinosaurs be "turbo-charged", but have fully ectothermic physiologies
Ontogenetic heterometabolism:
In most animals, metabolic rates slow down as age (and thus size) increases
Perhaps in dinosaurs was more extreme
Problem: No good living examples
Ancient Atmospheres
Even as dinosaurs were evolving, the atmosphere they were breathing was evolving, too. Geochemists have seen that the ratios of various gases, including oxygen, have varied over geologic time. At least some models suggest that the Middle Jurassic though the end of the Cretaceous had oxygen levels exceeding the present 20%. This would mean that every breath a dinosaur took would have more oxygen, making it easier to power a high metabolism.
Furthermore, experiments of growing plants of Mesozoic varieties under Mesozoic-style atmospheres suggests that their productivity (essentially, the amount of nutrients they produce per area per unit time) could go up 2 to 3 times present day conditions. If so, then there would have been more food available per unit area for the herbivores (and from this up the energy pyramid), again making it easier to be an endotherm in these conditions.
Warm-blooded Protocrocs?: Most studies assume that endothermy evolved sometime after the bird lineage (Ornithodira) and the crocodilian lineage (Pseudosuchia) diverged from each other. This is because crocodilians are ectotherms, as are all the next several outgroups (lepidosaurs, turtles). However, what if crocodilians were not ancestrally ectotherms, but instead reverted to a cold-blooded physiology from warm-blooded ancestors?
There is some evidence that this is the case:
Most extinct outgroups to the living crocodilians had a more upright stance (in fact, many of them parasagittal) allowing for more aerobic breathing (and at least one genus shows growth rates comparable to dinosaurs, pterosaurs, and mammals)
Crocs retain many ancestral features (unidirectional flow in the lungs; four chambered heart; etc.) that are useful for endotherms but not at all required for ectotherms (since lepidosaurs, turtles, and amphibians do fine without them)
Crocs have mitochondria of the "leaky" type used in endotherms to generate extra heat: however, they have far fewer of them per cell than living birds and mammals.
Living crocodilians are aquatic ambush predators living in thermally stable environments: these are conditions which would favor selection away from high metabolic rates to slower ones
This has led to speculation that the ancestral archosaurs were in fact more warm-blooded than crocodilians, and that the latter evolved "cooler blood" after the divergence of their lineage from other types of pseudosuchians. Thus, the origin of avian warm-bloodedness would not have occurred within Dinosauria, but at least in part before the bird line-croc line split.
A New(-ish) Idea: Mesothermy
In 2014 a study came out proposing that dinosaurs were intermediate between endotherms and ecotherms, and the authors termed them "mesotherms". (In fact, Dr. Scott Sampson had proposed the concept and the name "mesothermy" years earlier...). The particular study estimated both the maximum growth rate of fossil dinosaurs and their inferred metabolic rate (based in part on growth rate, so the whole study may wind up being a circular argument...). They found that most Mesozoic dinosaurs (including Archaeopteryx) fell in a range intermediate between where modern endotherms and modern ectotherms plotted (but in the same region as such animals as tunas, sharks, echidnas, etc.)
The authors interpreted this to mean that dinosaurs had the ability to generate internal heat, but did not greatly regulate their
body temperature. So in fact, what they call "mesothermy" is technically not intermediate between endothermy and ectothermy, but between homeothermy and poikiliothermy. And thus dinosaurs in their interpretation would be in terms of this course endothermic mesotherms. In their interpretation, the rise of actual warm-bloodedness in the bird lineage occurred somewhere well within Pygostylia. Future analyses will have to be done to see if this model is upheld.
Dinosaurs of the Middle Jurassic Xiashaximiao Formation of China: the sauropod Shunosaurus, the theropod Gasosaurus, and the stegosaur Huayangosaurus, by Gregory S. Paul




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}