Key Points:
•Modern animals are often characterized as "warm-blooded" (mammals, birds) and "cold-blooded" (everything else)
•This is a simplification of several related phenomena: energy source (endothermy vs. ectothermy); metabolic rate (tachymetabolism vs. bradymetabolism); and temperature stability (homeothermy vs. poikilothermy)
•Dinosaur species were initially inferred to be "cold-blooded", but similarity in posture and other traits led Owen to suggest they might have been warm-blooded; since that time various researchers have examined the alternatives.
•Dinosaur posture, locomotion, feeding adaptations, growth rates, bone texture, inferred respiration, and predator-prey ratios point to elevated metabolisms relative to today's non-avian sauropsids.
Pterosaurs
Not all reptiles of the Mesozoic were dinosaurs. There was a diversity of reptiles on land, and also in the sea and in the air.
A number of Permian & Triassic gliding reptiles, but only two powered flying groups:
Avialians (i.e., birds, a type of theropod dinosaur)
Pterosauria (pterosaurs, probably the sister group to
Dinosauromorpha within Ornithodira, but some consider them more primitive archosauriforms)
Pterosaurs:
First appear in Late Triassic; died out at very end of Cretaceous
Had simple hinge-like ankle and S-shaped neck of other ornithodirans, but perhaps not parasaggital stance
Skin stretched between finger and body, between legs, and from arm to neck
Wings reinforced by special internal fibres attached to muscles: could control shape of wing
Body covered with hair-like structures
Some used to think they walked bipedally, but most evidence suggests quadrupedal (almost gorilla-like), at least for the derived groups. (Primitive forms may not have been able to do much walking at all.)
Active powered fliers with large muscle attachments
Quite probably warm-blooded (insulated; very fast growth rates; active fliers)
Babies seem to have been able to fly right after hatching!
Earlier forms relatively small, short metacarpi, and long tails; later forms (Pterodactyloidea, the true pterodactyls) could be very large, had long metacarpi, and short tails and were better adapted at walking
Variety of diets: insectivores, fish eaters, fruit eaters, etc.
No evidence of direct competition between birds and pterosaurs.
Marine Sauropsids
Throughout Earth History, many reptiles (and other amniotes) have returned to the sea:
Abundant food
Equitable temperatures
Easy migrations
However, aquatic amniotes have to deal with:
Breathing (remain air-breathers)
Feeding (small fish, large fish & amniotes, shellfish, vegetation, etc.)
Locomotion (flippers, fins, etc.)
Reproduction (come out of water to lay eggs or some form of internalized reproduction)
First reptiles to return to an aquatic life were
mesosaurs:
Early Permian of Gondwana
Members of the primitive reptile group Parareptilia
Long needle-like teeth for catching small fish
Big (webbed?) hands and feet for paddling, tall deep tail for swimming
Could probably crawl on land, although probably not too well
Now known to have retained eggs (or young) in the body until they were developed enough to swim
Probably did not travel far from shore
Most primitive relatives of Mesozoic marine sauropsids were similar in general form (long needle-like teeth, webbed hands and feet, deep tail, some terrestrial ability, probably shore-dwelling or fresh-water) to mesosaurs, but later forms become more specialized for life in the sea.
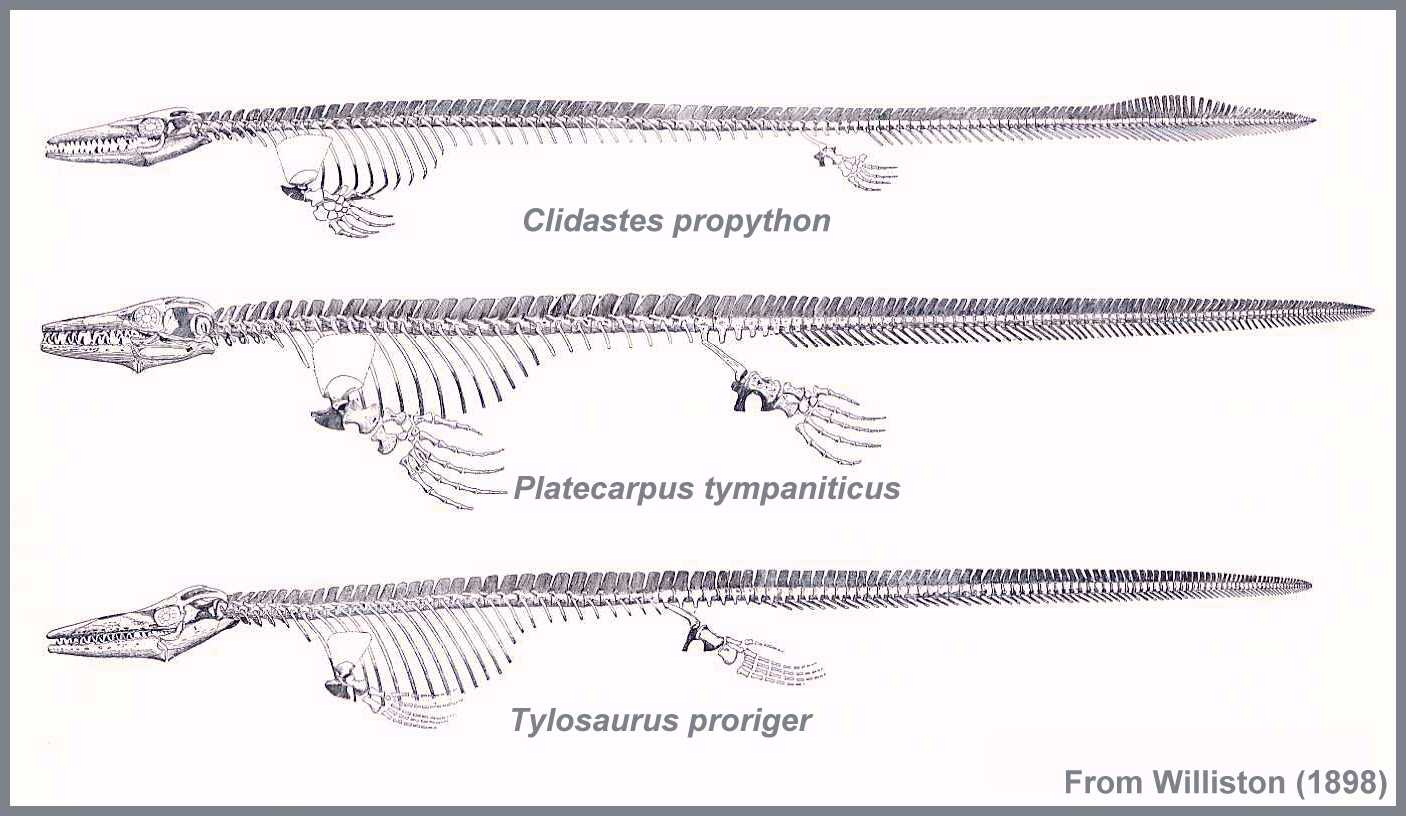
Many different clades of Mesozoic marine sauropsids, from almost every clade:
Testudines (marine turtles)
Lepidosauria, both Rhynchocephalia (pleurosaurs) and Squamata (mosasauroids)
Pseudosuchia (a small radiation of Middle Triassic marine poposauroids and two major types of marine crocodyliforms: the Jurassic-Early Cretaceous thallatosuchians and the Late Cretaceous-Paleogene dyrosaurs)
Even Dinosauria (hesperornithiform birds)
We will cover the most diverse and highly specialized forms: euryapsids (esp. ichthyosaurs and plesiosaurs), mosasaurs, and marine turtles.
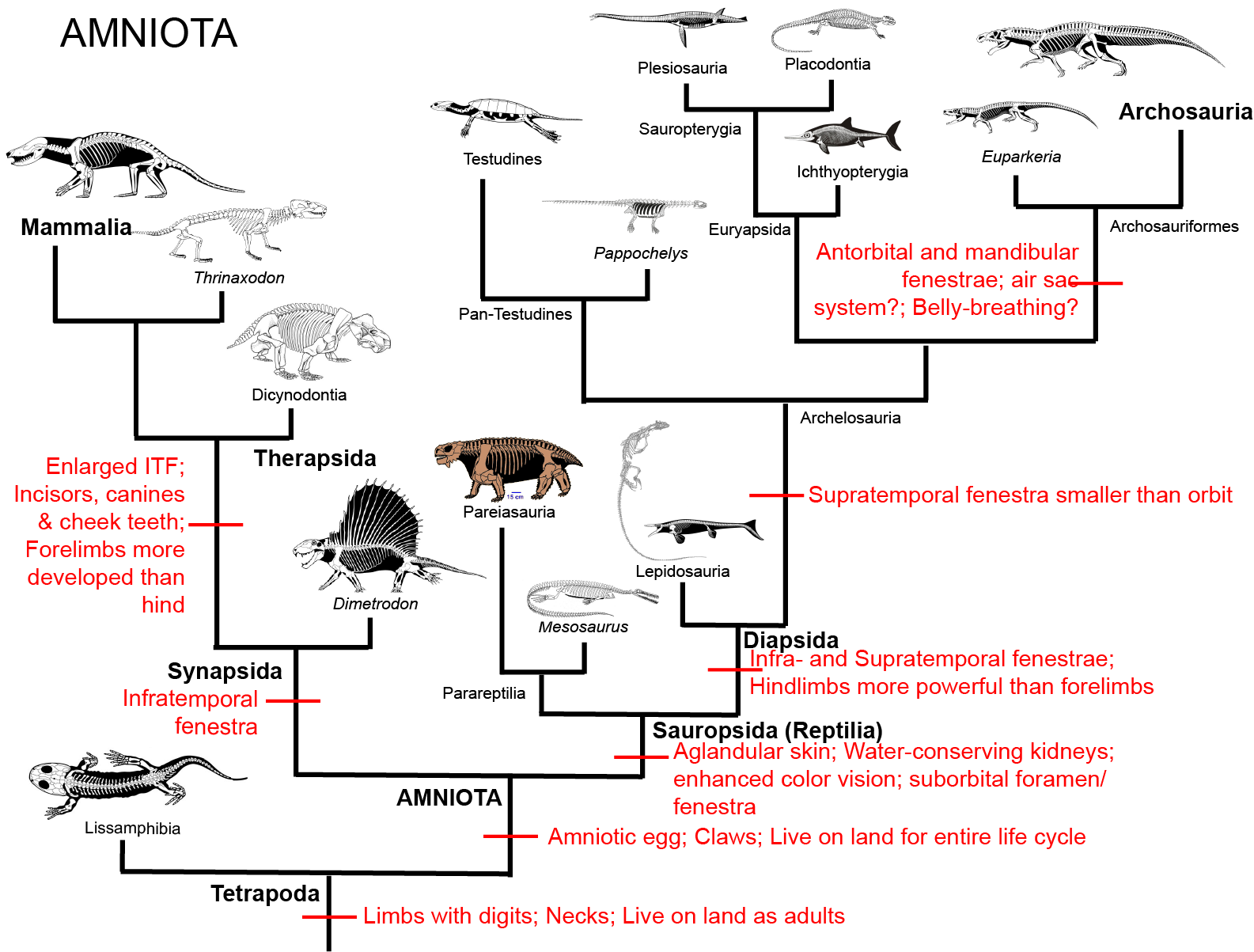
A reminder of amniote phylogeny: :
Euryapsida: more closely related to Archosauria than to Lepidosauria, so part of the larger clade of Archosauromorpha. Euryapsids:
First appear in Early Triassic
They MIGHT be basal members of the lineage that contains turtles, if turtles turn out to be archosauromorphs
Most primitive members have webbed fingers and may have crawled around on shore
Early euryapsids (and most later ones) seem to have been fish eaters, with needle- or cone-shaped teeth.
Remains show that even primitive euryapsids retained young inside body
until birth, thus allowing them to live their entire life in the water. This allowed for the extreme specializations of the two
highly derived pelagic (open seas) euryapsid groups: ichthyosaurs and plesiosaurs
Most of long snouts and cone-shaped teeth: fish or squid-eaters for the most part. However, some early forms had short snouts and no teeth,
and may have been
suction feeders
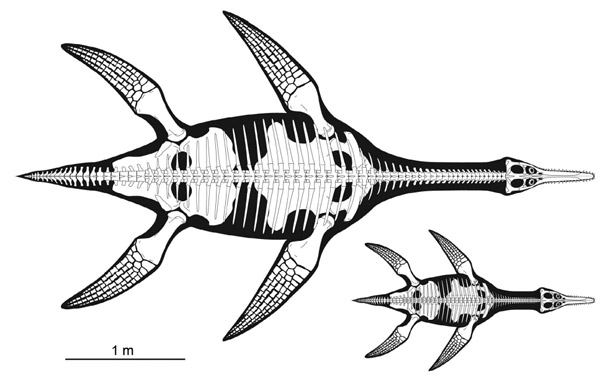
First appear in latest Triassic; die out at the end of the Cretaceous
Fore- and hindlimbs turned into large flippers for swimming; tail generally short
Two general body types:
Small head, needle-like teeth, long neck: small fish or squid eaters
Large head, big cone-shaped teeth, shorter neck: large fish or marine reptile eater
The large-headed forms appear to evolve a number of times from the long-necks
Include the largest known marine reptiles of all (bigger than all theropods, and in
fact bigger than all dinosaurs other than sauropods!)
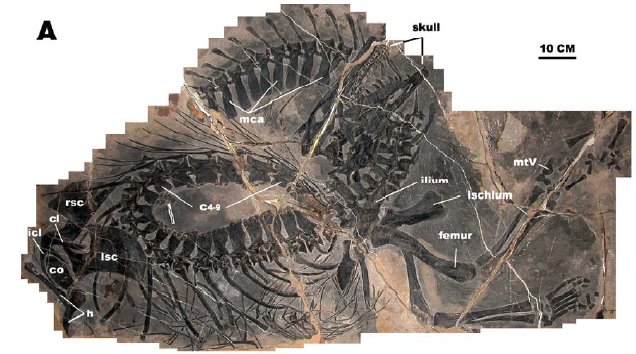
A newly-described fossil shows that at least some plesiosaurs gave birth live to a
single large-bodied baby (more like whales than the many small babies found in ichthyosaurs, mosasaurs, and sea snakes). This may in fact
suggest extended parental care for at least the derived plesiosaurs.
A couple of different closely-related families of marine turtles
First groups are late Early Cretaceous; some died out in Early Cenozoic, others survive to today
Fore- and hindlimbs turned to flippers, forelimbs provide must of the thrust
Feed on shellfish, fish, jellyfish, vegetation, etc.
A newly discovered form actually used suction feeding
Shells are typically thinner than land-living turtles
Crawl onto beach to lay eggs, but otherwise fully marine
Recent work shows that the pelagic ichthyosaurs and plesiosaurs (and possibly the mosasaurs) had an elevated metabolism, more like endotherms
than ectotherms.
Origin of the Fuzzballs: The First 7/10ths of Mammalian History
On land during the Mesozoic, there were plenty of organisms other than dinosaurs. Among the most important (especially for us!) were the mammals.
Mammals and their closest relatives (more properly Mammaliformes, or sometimes "Mammaliaformes") appear in fossil record the same time as dinosaurs, in
Late Triassic.
Mammals are very advanced therapsids synapsids.
True mammals (Mammalia) found from Late Triassic onward.
Like birds, many of the features that characterize modern mammals don't fossilize:
Warm-blooded
Covered with fur
Sweat glands
Mammary glands
Parental care of young
On the other hand, some mammalian features are preservable:
Only two sets of teeth: deciduous ("baby") and permanent
Highly specialized teeth divided into incisors, canines, premolars, and molars
Teeth highly distinctive, can recognize a species from only one or two cheek teeth (premolars and molars)
Lower jaw comprised only of dentary; jaw joint is dentary-squamosal, not
quadrate-articular
However, this trait turns out to be convergent between the living groups, and not present in all fossil mammals
Respiration using diaphragm (so that dorsal vertebrae are divided into thoracic and lumbar sections)
And more
Many features limited to Mammalia among living amniotes were probably found in their closest non-mammalian therapsids relatives. For example, we can't say for certan when warm-blood, fur, sweat & mammary glands show up. We can determine a few of these, though:
Evidence for parental care in basal synapsids, as well as more derived therapsids
Division of teeth into incisiors, canines, and cheek teeth in early therapsids
Diaphragm breating in more advanced therapsids
Predator-prey ratios of therapsid communities suggest elevated (or at least non-cold blooded) metabolisms
No direct evidence of fur outside Mammaliformes, but a likely osteological correlate to whiskers and a mobile wet nose are present outside Mammaliformes. (Developmentally these are also linked to mammary glands, too!) More primitive versions of fur may go down much further in synapsid history, but we don't yet have fossils to constrain this.
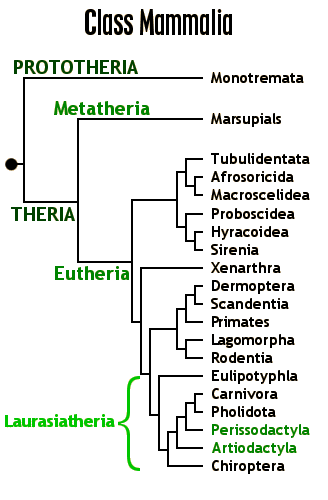
Monotremes: the last surviving egg-laying mammals. Rare and surviving only in Australasia
Marsupials: reproduce by young born live but very poorly developed, which then suckle and grow within a pouch. Dominate Australasia, common in South America, present in early Cenozoic Europe and North America
Placentals: stay in the womb until more developed, fed by a placenta. The dominant group in the Americas, Africa, and Eurasia.
However, mammal diversity in the Mesozoic was MUCH different.
Many groups of Mesozoic mammals have long since died out. And some Mesozoic mammal groups survived the end of the Cretaceous, but have since died out.
Most Mesozoic mammals very small (shrew-to-house cat sized, with a few badger-sizedforms in the Cretaceous); mammals only become large AFTER extinction of non-avian dinosaurs.
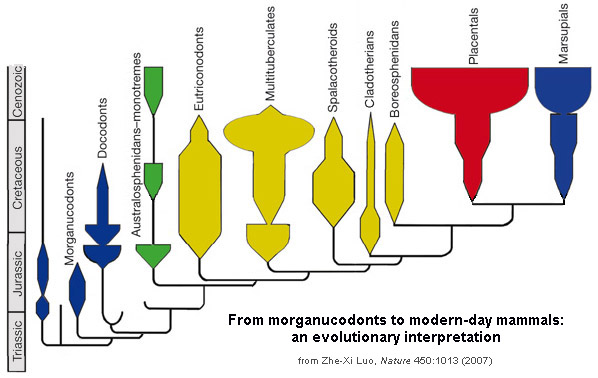
Some major groups of Jurassic and Cretaceous mammals:
Prototheria (sometimes called "Australosphenida"; monotremes and their extinct relatives):
Generally relatively stumpy legs and tails, even in modern forms
Oldest from Middle Jurassic onward
Oldest monotremes proper from Early Cretaceous (perhaps actually a platypus! it has the same electrosensory apparatus); survive today in Australasia as platypus and echidna
Prototheres were never a dominant group, but were once moderately common in Gondwana (present in Asia, too)
Retain ancestral sprawlling posture and egg-laying reproduction
In their time, were major parts of the larger mammal fauna: ecologically comparable to opposums and raccoons in the modern U.S.
Wiped out at end of Cretaceous
Mode of reproduction unknown
Allotheria (allotheres):
Some studies suggest that the basal, poorly known Late Triassic "haramiyids" (a paraphyletic group) and the Jurassic tree-climbing euharamiyids are part of this clade; others put these branches outside the Prototherian + Allotheria + Theria group
More definite members are the Cretaceous Gondwanatheria, and the diverse Multituberculata.
Gondwanatheria
Until recently known only from isolated jaws and teeth, which indicated a gnawing (rodent-like) diet
As the name implies, known only from the southern continents
New discovery of nearly complete skull of Vintana of the Late Cretaceous of Madagascar shows at least some gondwanatheres had relatively big brains, very large eyes, and well-developed olfaction. Also, Vintana is quite large (about beaver-sized), making it among the largest of Mesozoic mammals
Some gondwanatheres survived in South America until the Miocene Epoch (a mere 18 million years ago!)
At least some studies place Gondwanatheria WITHIN Multituberculata
Multituberculata
Oldest multituberculate fossils Middle Jurassic; survived into early Cenozoic (about 35 Ma) when they became extinct
Specialized molars and gnawing teeth: convergent with rodents, and a major radiation in this way of life before the true rodents evolved
Recently discovered trace fossils shows that some of these gnawed on dinosaur bones!
There are other branches of early mammals (docodonts, symmetrodonts, dryolestoids, etc.), but the most important remaining two are joined together as the clade Theria. Therians are united by various skeletal (parasaggital stance, some dental, etc.) and soft-tissue (nipples, external ears, etc.) features. Therians include the
metatheres and eutheres, which diverged by the Early Cretaceous. (Supposed Jurassic eutherian Juramaia now appears to be outside of Theria proper).
Metatheria (marsupials and their extinct relatives):
Recent phylogenetic analyses show that there are no definite members of Marsupalia during the Mesozoic, although they are present so early
in the Cenozoic that they probably had evolved before the end of the Cretaceous
Eutheria (placentals and our extinct relatives):
The earliest Late Jurassic Juramaia and the Early Cretaceous Eomaia were once thought to be eutheres. However, new analyses place these two outside Theria. As a result, the oldest known eutherians presently are from the Late Cretaceous, but the clade must extend down into the Early Cretaceous.
Survive today as most diverse group of mammals (including us!)
Placental mammals reproduce by keeping young in womb until birth, fed by placenta: not certain how non-placental eutheres reproduced
Mesozoic eutheres were small; many were herbivorous, omnivorous, and insectivorous
True placentals are not yet known older than the end of the Cretaceous (in fact, the most recent comprehensive study shows that none of the Cretaceous eutheres definitely belong to Placentalia),
but it is quite possible that the major divergences had already happened before the end of the Age of Dinosaurs
Prototheres, allotheres (as multitubercultates), metatheres (including the first marsupials), and eutheres (including the first placentals) all survived the great extinction event at the end of the Cretaceous.
The Flowering of the Age of Dinosaurs: Plants of the Mesozoic
The base of the food chain on land is plants. They are responsible for
taking sunlight, carbon dioxide, and water and combining them to produce glucose and oxygen (in other words, the reverse of the aerobic
respiration equation).
Plants at the dawn of the age of dinosaurs were very different
from those of the modern world: not so much of what was there as what wasn't. There was no grass, no grain, no fruit, no flowers. But by the end
of the Mesozoic, these were present.
Primitive plants are spore plants. Spore plants are a paraphyletic grade: some are more closely related to seed plants than are others. Spore
plants first colonized land in the early Paleozoic Era.
They reproduce by releasing
spores, which settle onto moist surfaces, grow into plants that produce sex cells which meet in the thin film of water, and join to produce
a new spore-producing plant. (Note that this is somewhat analogous to amphibian-grade tetrapods: plants that live their life on land, but need
to put their sex cells in water to reproduce.)
All of these were present in the Mesozoic. For most of the Mesozoic the dominant ground cover was ferns, and tree ferns were fairly important
trees in the Triassic and Jurassic.
Spore plants lack true wood (tree ferns cheat by having many stalks growing right next to each other for support), nor do they have complex root
systems.
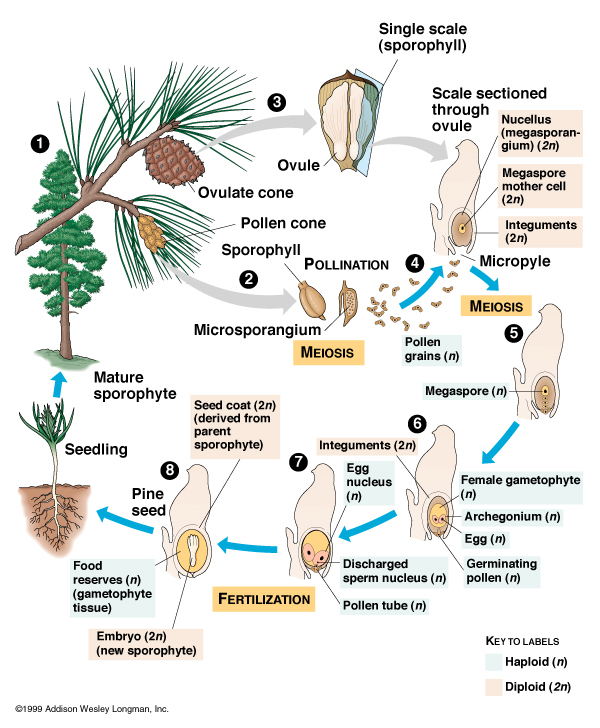
Those traits, however, ARE present in the seed plants. Seed plants first appear
in the mid-Paleozoic Era, and become the dominant land plants in the Permian Period of the Paleozoic Era.
They reproduce by releasing male sex cells (carried in pollen) which land on
female sex organs, join with female sex cells, produce a fertilized seed, which can then be released from the plant to land in the soil and
germinate on its own. (This is analogous to the amniotic egg in tetrapods, allowing plants to colonize further inland into drier regions.)
All non-flowering seed plants are traditionally grouped into a paraphyletic grade, the "gymnosperms": some extinct gymnosperms are more closely related
to flowering plants than to other gymnosperms. The living gymnosperms do form a clade (Acrogymnospermae). Various sorts of gymnosperms exist:
Cycads, once widespread, but now more common in the tropical regions. They
look something like palms and something like pineapples, but are neither. The include small tree- and shrub-sized species. They were VERY common in the Mesozoic.
Ginkgoes, trees that were widespread in the Mesozoic
and Cenozoic but now limited to a single species
Conifers, highly diverse tree-and-shrub sized species including:
Bennettitaleans, including
tree and shrub forms and the only fully extinct group listed in this page!
As today, gymnosperms were an important group of land plants in the Mesozoic. In fact, they were even more common then! Ginkgoes, dawn redwoods, cycads, and bennettitaleans (all rare or extinct today) were major parts of the flora, and major sorts of dinosaur food.
Some gymnosperms (conifers, cycads) wrap their seeds in a fleshy coating, and some (bennitataleans, cycads) had specialized structures around their female sex organs to attract insects. But only in the next group do we get true fruit and true flowers.
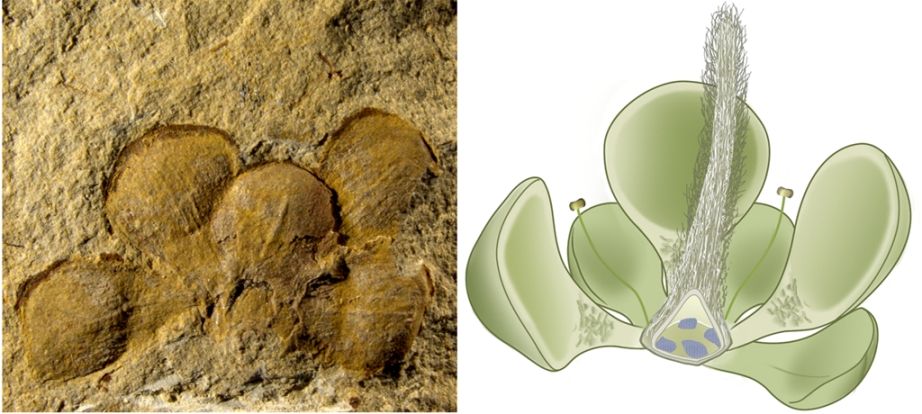
Flowering plants, called the angiosperms or anthophytes are the major clade of modern plants. With rare exceptions, if you have eaten a plant, it was an angiosperm. Angiosperms are a clade within the seed plants. Their mode of reproduction is to develop a specialized set of both male and female sex organs within a flower; pollinators are lured to the flower, pick up pollen, have pollen rub off on the flower of another plant, where they fertilize the female sex cells, and a seed is made. That seed is covered by a coating
of fleshy or nutty tissue: the fruit.
The basic angiosperm life cycle hinges on co-evolution with animals:
Bright colors, attractive smells, and interesting patterns on the flowers attract pollinators
(typically flying insects). These move pollen (containing the male sex cells) to flowers of other plants (where the female sex cells are)
Fruit remains bitter, hard, and dull colored until the seeds are ready to grow. At that point, the fruit becomes brightly colored, fleshy or nutty, and sweet and juicy. The fruit is then eaten by a vertebrate, which leaves the area and deposits the seeds in its dung.
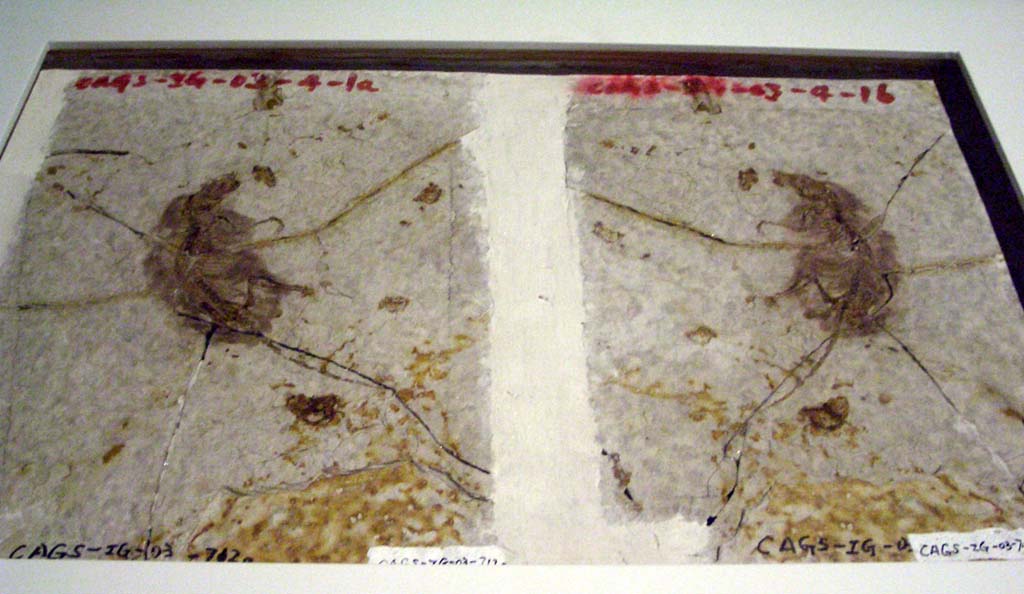
Possible angiospermbody fossils are known from the Jurassic, and close relatives of the angiosperms go back to the Permian, but the oldest definite angiosperms are from the Early Cretaceous. Early Cretaceous angiosperm pollen and leaves are known from far off Prince George's County, Maryland and nearby in Virginia, and similar fossils are known from earlier in the Cretaceous in China. Pollen of angiosperms (or near-angiosperms) date back to the Middle Triassic, but we can't tell yet if these are from true flowering plants or pre-flowering relatives of the angiosperms.
If angiosperms evolved flowers and fruit in the Cretaceous, who were their target audiences?
Flower targets?
Earliest Cretaceous "birds" (flying theropods) were still relatively rare, and none show nectar-eating adaptations.
Mammals were small, and may have been pollinators, but no bats (the main modern mammalian pollinators) yet.
Insects (including beetles, flies, lepiodopterans, and hymenopterans) were present, and were almost certainly the main pollinators in the Mesozoic just as they are today.
Fruit targets?
Herbivorous mammals of the Early Cretaceous were small, and may have eaten berry-sized fruit, but no more than that
Hebivorous crocodyliforms were rare
But herbivorous dinosaurs were VERY common, and could easily eat and transport a LOT of fruit!
So thank the insects for flowers, and thank the dinosaurs for fruit.
The rise of the angiosperms occurs about the same time that low-browsing herbivorous dinosaurs (ankylosaurs, iguanodontians, rebbachisaurids) become dominant over medium (stegosaur) and high (typical sauropod) browsers. Are these changes linked? Although angiosperms were present in the Early Cretaceous, they seem to have been relatively rare then, and unlikely to have been a major food source for these groups at first. But it may be that increase in low-browsing forms favored the spread of herb-sized angiosperms.
By the Late Cretaceous many modern clades of angiosperms were present (mangolias, rose-relatives, maples, etc.). Also during this time the first grass appears. Grasses include not only the stuff that grows in lawns and meadows, but all the grain-producing plants (wheat, barley, etc.), as well as bamboo. Their flowers are extremely small, and they are often wind-pollinated rather than by the help of insects.
Grasses grow from the base of the leaf rather than the tip. They often have little bits of silica in them to persuade herbivores not to eat them. Recent discoveries in both Laurasia and Gondwana: the latter were found in titanosaur sauropod coprolites! So at least some Cretaceous dinosaurs were grass eaters. However, grasses seem to have been relatively rare in the Mesozoic, and did not form grasslands until much later. Ground cover in the later Mesozoic was a mixture of ferns and herbaceous angiosperms. So as far as we know, no dinosaur other than birds ever wandered in prairies or savannas: these appear much later, long after the end of the Mesozoic.




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}