
Toward Amniota
The Reptiliomorph Stem
Before we start: As we advance up the reptiliomorph stem, a vital transformation occurs that we can only indirectly infer, but must always remember: The evolution of the cleidoic (aka amniotic) egg:
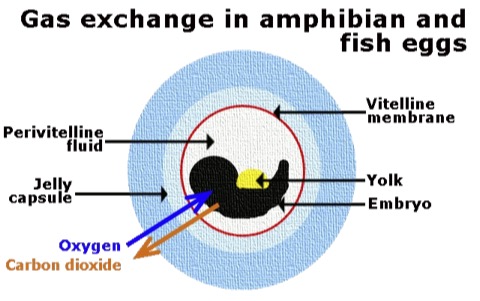
- Anamniotic egg: The ancestral state for Choanata (at least). Embryo and yolk enclosed inside vitelline membrane surrounded by a jelly capsule. The reproductive strategy at hand for basal reptiliomorphs, but:
- The egg is vulnerable to desiccation so must be laid in a moist environment - typically a body of water (needn't be large).
- Typically laid in environment full of predators
- Embryonic development is slow, limited by the rate of simple diffusion of oxygen and carbon dioxide.
- Neonate, upon hatching, must be water-breathing.

- Cleidoic egg: Appearing somewhere below the base of Amniota. It is air-breathing, meaning that it can be laid on land (in the Carboniferous, that meant "away from the scariest predators." But it required many derived features to work together, both in the egg, itself, and in the adults who made them.
-
Extraembryonic membranes (outgrowths of the embryo) facilitate accelerated metabolism and growth:
- Amnion: encloses embryo directly, providing aqueous environment
- Chorion: facilitates gas exchange with air, encloses amnion, yolk, and allantois.
- Allantois: facilitates waste elimination
- albumen, which provides a reservoir of water and proteins, and protects against pathogens,
- a shell (ancestrally leathery) that separates interior and external environment.
- To make a shelled egg meant fertilizing it before the shell was secreted, thus internal fertilization was required. In amniotes, this is accomplished by means of a penis. (Other critters use different intromittant organs.)
- To lay them safely usually required burying them. To that end, amniotes have claws for scratch-digging.
- Getting out of the egg required an egg-tooth at the tip of the neonate's snout.
- Of course, the neonate needed to be fully air-breathing.
Synapomorphies and trends:
- Gastrocentrous vertebrae: with large pleurocentra, often coossified with neural arches, and small crescent-shaped intercentra.
- As in anthracosaurs, early members of the reptiliomorph stem display a contact between tabulars and parietals. Whether this is a synapomorphy of reptiliomorpha or a larger group is not clear.
- Recall that there is no indication that any reptiliomorph retained gills as a metamorphosed adult.
Reptiliomorph diversity:
Gephyrostegida: (Late Carboniferous) Relatively small, (Gephyrostegus has 22 cm snout-vent. Length). Gephyrostegus has the general proportions of a land animal with these reservations:- Its head is relatively large
- Its vertebrae are not strongly coossified. (Carroll, 1970)
Larvae unknown.
Chroniosuchia: (Middle Permian - Late Triassic) By Late Triassic, these were the last surviving non-amniote reptiliomorphs. Distinctive features include:- armor of interlocking scutes on their backs.
- Snouts perforated by antorbital fenestrae.
Hypotheses of phylogeny place them all over the tree, from a position outside crown group Tetrapoda to one close to Amniota. All authors agree that they are not on the amphibian branch.
Larvae unknown.
Seymouriamorpha: (Latest Carboniferous - Permian) Small animals with plesiomorphic aquatic larvae but adults ranging from the terrestrial Seymouria to paedomorphic aquatic forms. (Note: this is the last clear occurrence of water-breathing larvae on the reptiliomorph stem.)
Seymouria baylorensis (Permian)- Stocky bodies and short tails, indicative of adult life on land
- Impedance-matching ears in deep embayments at the rear of the skull. The fenestra ovalis is enclosed in a long tube formed from the basisphenoid and parsphenoid - a condition unique to Seymouriamorpha. Despite this transformation of the auditory system, the labyrinth organ and brain of Seymour appear plesiomorphic. (Bazzana-Adams et al., 2023)
- Swollen neural arches. (Note: this is also seen in many basal amniotes. Convergence?)
- Very large adults of the terrestrial forms like Seymouria incorporate a second sacral rib pair and lose sensory canals in skull.
And yet, they present striking plesiomorphies:
- Discosaurus: retains the anocleithrum above its cleithrum.

And now, a mess:
Traditional paleontology has recognized Lepospondyli: (Carboniferous - Permian) An amazingly diverse group of small tetrapods characterized by trends toward:- Holospondylous vertabrae in which the pleurocentrum is fused to the neural arch. (Some basal forms have small intercentra.)
- Loss of skull bones including the supratemporal and intertemporal.
- Paired occipital condyles (convergent with Temnospondyli. Link to Pytonius.)
- "Microsaurs" (Late Carboniferous - Early Permian) - a grade group of small tetrapods including Recumbirostra, a clade (probably) containing long-bodied, short limbed or limbless creatures modified for fossorial or aquatic life.
- Nectridea (Late Carboniferous - Late Permian) - strongly aquatic creatures including the pop-culture favorite Diplocaulus.
- Aistopoda (Early Carboniferous - Early Permian) - Limbless aquatic eel-like forms that appear surprisingly early for stem amniotes.
The lepospondyl death-watch: But it appears that Lepospondyli is not a thing. Work of the last two decades has eviscerated it with:
- Aistopoda being convincingly demoted to the tetrapod stem. (Pardo et al., 2017) See the last lecture.
- Recumbirostran "microsaurs" being booted upward toward Amniota. (Jenkins et al., 2025)
Former lepospondyl groups:- Non-recumbirostran "Microsauria": (Mid- late Carboniferous) The evisceration of traditional "Microsauria" by the removal of Recumbirostra leaves a few remaining taxa including:
- Utaherpeton franklini: (mid Carboniferous) The earliest traditional "microsaur," and phylogenetically basal. Salamander size and shape (snout-vent length 4 cm).
- Tuditanidae: (Late Carboniferous) Superficially reptile-like with proportionally larger limbs and jaw joint posterior to occipital condyle. Thought to be ecologically similar to captorhinids. (See below.)
- "Nectridea:" (Carboniferous - Permian) Specialized aquatic forms with:
- Haemal arches that fuse directly to pleurocentra.
Nectridia includes Diplocaulidae (E.G. Diplocaulus magnicornis (right and link to full skeleton). Characterized by:- Bizarre expansion of the posterolateral corners of the skull table into conspicuous "horns."
- Convergent evolution of wide interpterygoid vacuities.
With their large interpterygoid vacuities and sutured basipterygoid articulations, these are reminiscent of stereospondyl temnospondyls. I'm not claiming anything except that nectrideans are overdue for a reexamination.
- Solenodonsaurus: (Late Carboniferous) Comparatively largish (45 cm snout-vent length) tetrapod with adaptations for terrestrial life including humerus with a slender shaft, triangular head unsuitable for aquatic feeding, and complete lack of bony lateral line canals. Unlike terrestrial seymouriamorphs, the otic notch is small and lacks evidence for a tympanum.
For most of the history of its study, it has been considered important to understanding the rise of proper amniotes, however it has not been considered one since its first cladistics analysis by Gauthier et al., 1988.. Later, Danto et al., 2012 found Solenodonsaurus to be the sister taxon of "Lepospondyl," but as Lepospondyli has fallen apart, its position needs to be reassessed.
- Westlothiana: (Early Carboniferous) The earliest known unambiguous tetrapod yet surprisingly far up the amniote stem. In contrast to Solenodonsaurus a mere 10 cm in snout-vent length. Elongate with small limbs, it is reminiscent of some "microsaurs" or of many living fossorial skinks. Because it fits with notion that the first cleidoic eggs were laid by small creatures, Westlothiana was at first hyped as "the first amniote," however phylogenetic analysis didn't bear this out. Instead it emerged as the sister taxon of "Lepospondyl."(Smithson et al., 2011) Without this anchor, its position is unclear.
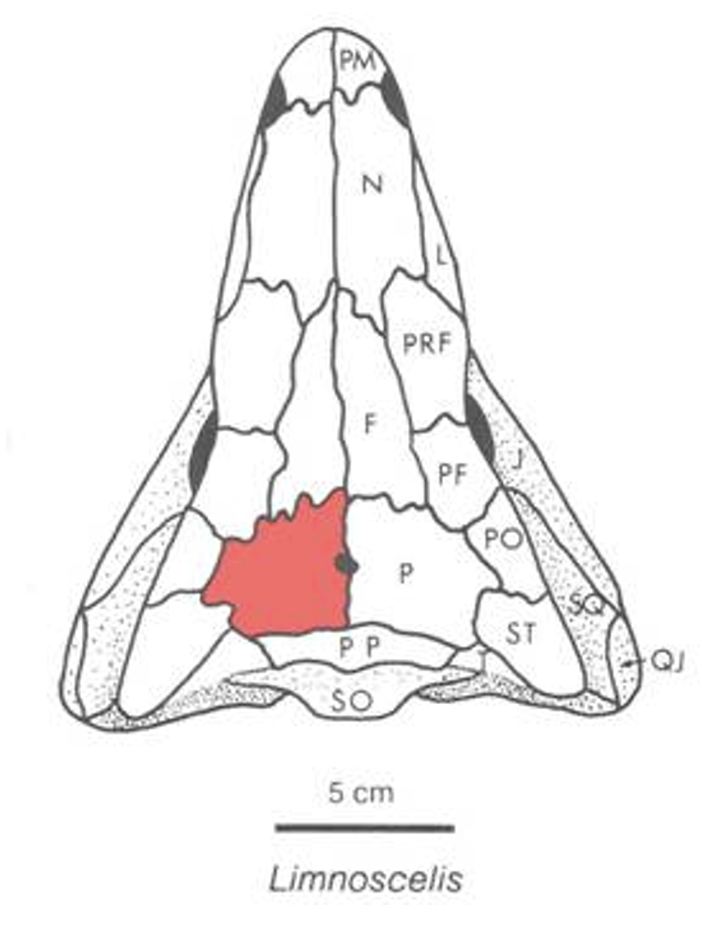
Limnoscelis skull © Stuart Sumida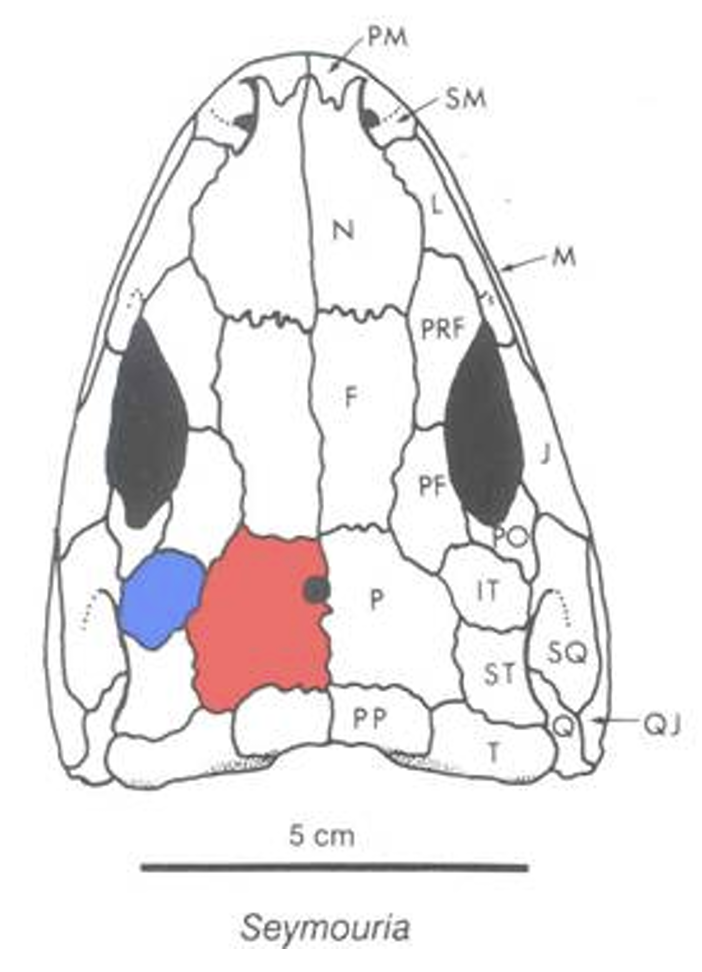
Seymouria skull © Stuart Sumida
Westlothiana is interesting because its well-preserved skull displays shared features with more derived stem amniotes like Limnoscelis (cf. plesiomorphic Seymouria) including:- Postparietals wide compared to tabulars.
- Postorbital contacts supratemporal, excluding parietal from squamosal
- Casineria: Another Early Carboniferous little guy, but this time, headless. Because of its fragmentary remains, its precise position is unclear, but it has one very interesting feature - its digits end apparently ended in claws. This is new, and considered a necessary prerequisite for the evolution of the cleidoic egg, whose parents needed to be able to scratch-dig holes in which to bury them. Did Casineria lay a fully cleidoic egg? GORK. Claws could have evolved for another purpose then been exalted for scratch-digging, but at least it could, in principle, bury any egg that it laid.
Emerging from "the Mess"
Recumbirostra: (Late Carboniferous - Early Permian) This ecologically diverse variety of small critters is the big news of the last decade. Previously deemed to be the monophyletic core of "Microsauria," recumbirostrans have been kicked up the amniote stem, with some members having been considered close to Captorhinidae (wait for it)(Pardo et al., 2017) at the base of Eureptilia (i.e. inside proper Amniota). But like Recumbirostra, the position of Captorhinidae has also been labile. Now, the features uniting the two clades are seen as plesiomorphies, and recent analyses have recovered both on the amniote stem. (Jenkins et al., 2025)
Recumbirostrans range from basal stout compact forms to derived long-bodied forms with small limbs. Older interpretations often regarded them as semi-aquatic, but more recent studies show them primarily to be a fossorial radiation, showing, to varying degrees, anatomic specializations for digging with the head. (Cf. living caecilians and amphisbaenian squamates.) Their synapomorphies relate to this ecology.
Synapomorphies of Recumbirostra:
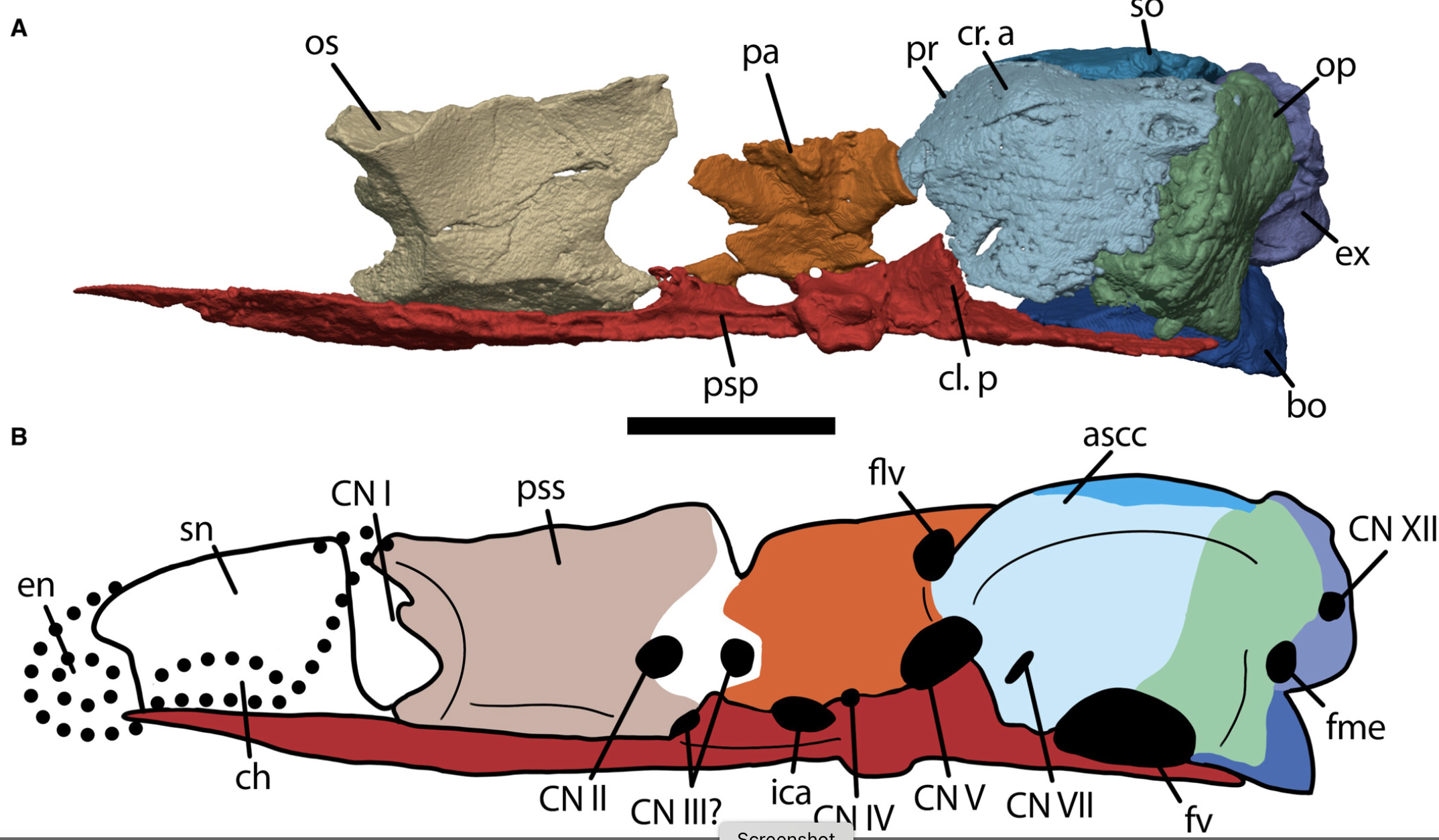
- Skull reinforced by strong ossifications of the pile antotica as pleurosphenoids and by paired orbitosphenoids in the sphenoid region.
- Stapes enlarged to cover fenestra vestibuli. Its shaft is stout and articulates with quadrate.
Recumbirostran diversity: Not entirely settled, but Jenkins et al., 2025 provide the last word, recognizing Pantylus (pictured above) as basal branch, followed by a dichotomy:
- Chthonosauria: Less strongly specialized for fossoriality, including Hapsidopareion, (pictured above), and the strangely derived Brachydectes. (only recently recognized as a recumbirostran.)
- Nastrondormes: Including more highly derived fossorial forms like Rhynchonkos.
Synapomorphies of Recumbirostra and Amniota:
- Supraoccipital that forms from a single ossification center and contributes to medial margin of otic capsule
- Absence of paired fossae of endolymphatic sack.

The transverse flange of the pterygoid in assorted amniotes plus the recumbirostran
Odonterpeton triangular (H) but not Rhynchonkos (I) from Mann et al., 2022- Transverse flange of pterygoid. This feature serves as the origin of the medial pterygoideus muscle, jaw adductors whose action is roughly perpendicular to that of the temporalis muscles originating on the braincase and skull roof. Its presence indicates the ability to maintain constant bite force regardless of the position of the jaw - a feature that enhances feeding strategies. But watch out! Porro et al., 2024 identify a transverse process in the pterygoid of the embolomere Eoherpeton, so we may have a poor understanding of this feature's distribution.
Cotylosauria
Cotylosauria (Late Carboniferous - Quaternary) is the LCA of amniotes and diadectomorphs and all descendants. Synapomorphies include:
- Complete absence of lateral line grooves
- Postparietal and tabular restricted to occipital surface.
- Sacrum with two sacral vertebrae
- Epidermal scales present (see below)
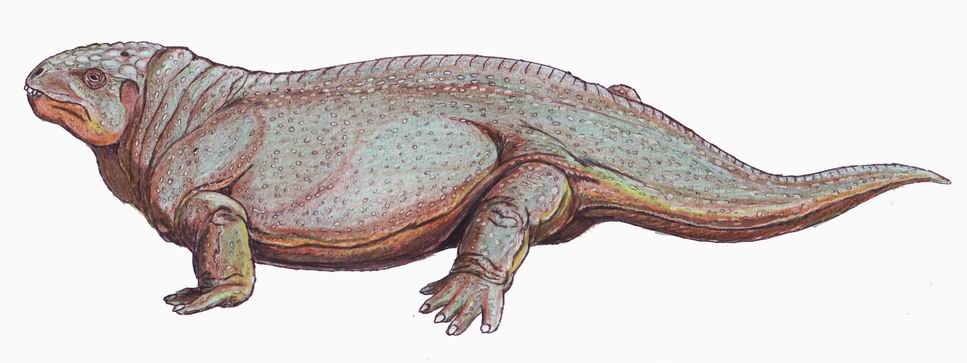
Diadectomorpha: (Late Carboniferous - Late Permian) includes the largest non-aquatic predator (Limnoscelis and the phylogenetically most basal herbivores (Diadectes and kin.) Well-known members include:
- Limnoscelis: (Late Carboniferous) At ~1 m snout-vent length, the apex terrestrial predator of its environment.
- Tseajaia: (Early Permian) Roughly half the size of Limnoscelis, a generalized predator.
- Orobates: (Early Permian) Iguana-sized with robust skull and blunt teeth suitable for herbivory.
- Diadectes: (Early - Late Permian) A large (1.5 m. snout-vent length) barrel-chested herbivore.
Synapomorphies of Diadectomorpha:
- Loss of intertemporal. (Incorporated into parietal as a lateral wing of parietal.)
- Supraoccipital extends beyond margin of otic capsule.
- Specialized articulation of atlas/axis complex with skull.
- Extremely robust adductor crest of ventral surface of the femur.
Limnoscelis skull © Stuart Sumida
Seymouria skull © Stuart Sumida
Points of interest:
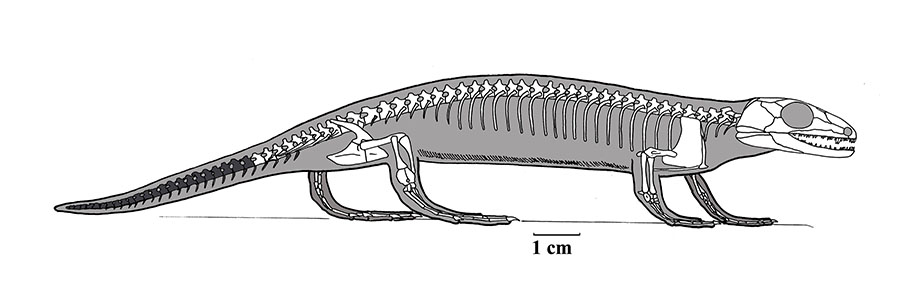
OroBOT - the robot OrobatesLocomotion: Orobates co-occurs with the ichnotaxon Ichniotherium sphaerodactylum. Because Orobates is known from a nearly complete skeleton, Nyakatura et al., 2019 were able to model its locomotion based on digital models developed from CT scans. These results were ground-truthed using a robotic replica.
The upshot: The limbs shouldered much more of the locomotor burden than in more basal reptiliomorphs. Although undulation of its trunk contributed to its locomotion, Orobates seems to have employed a semi-improved gait approaching that of crocodylians.
Epidermal scales?: The scales of amniotes are fundamentally different from those of fish, stem tetrapods, and basal stem amniotes. Those are derived from the ancestral dermal cosmoid scales of basal sarcopterygians. They persist up the reptiliomorph stem to the level of Solenodonsaurus and Westlothiana, and in the form of gastralia into Amniota. (Yes, Virginia, gastralia are a derived form of fish scales.) The scales of amniotes and their derivatives are epidermal, in contrast, and formed from keratin A and Keratin B - tough tissues but not bony. (Don't get confused! Many amniotes also have osteoderms, bony nodules arising from the dermis that have a deep developmental homology with fish scales, but are not directly homologous.
The origin of epidermal scales is mysterious, but Voigt et al., 2024 describe the convincing impressions of epidermal scales in the footprint ichnotaxon Ichniotherium cottae, which is attributed to a diadectid. For actual body fossils, see below.
Herbivory: Diadectomorphs include the very first creatures covered in this course that were primarily herbivorous. Evidence for this in Diadectes includes:
- Blunt robust incisor-like teeth in front
- Transversely wide multicusped cheek teeth
- Enlarged gut cavity
But why? The rise of Diadectomorphs coincides with the collapse of the classic Carboniferous coal swamp biome, perhaps providing an ecological opportunity. But to be an herbivore, one must be able to eat plant material. Ponstein et al., 2024 propose that the shift to herbivory was made possible by a change in lung ventilation.
- Lissamphibians use a buccal pump to ventilate the lungs, employing the ventral branchial and hyoid skeleton to pump air from the oral chamber to the lungs. This represents the plesiomorphic state for Tetrapoda.
- Amniotes use costal ventilation, elevating and lowering the ribs to expand that thoracic cavity, drawing air into the lungs.
But note: A minority view holds that Diadectomorphs are amniotes - Sister taxon to Synapsida. (Ponstein et al., 2024.)

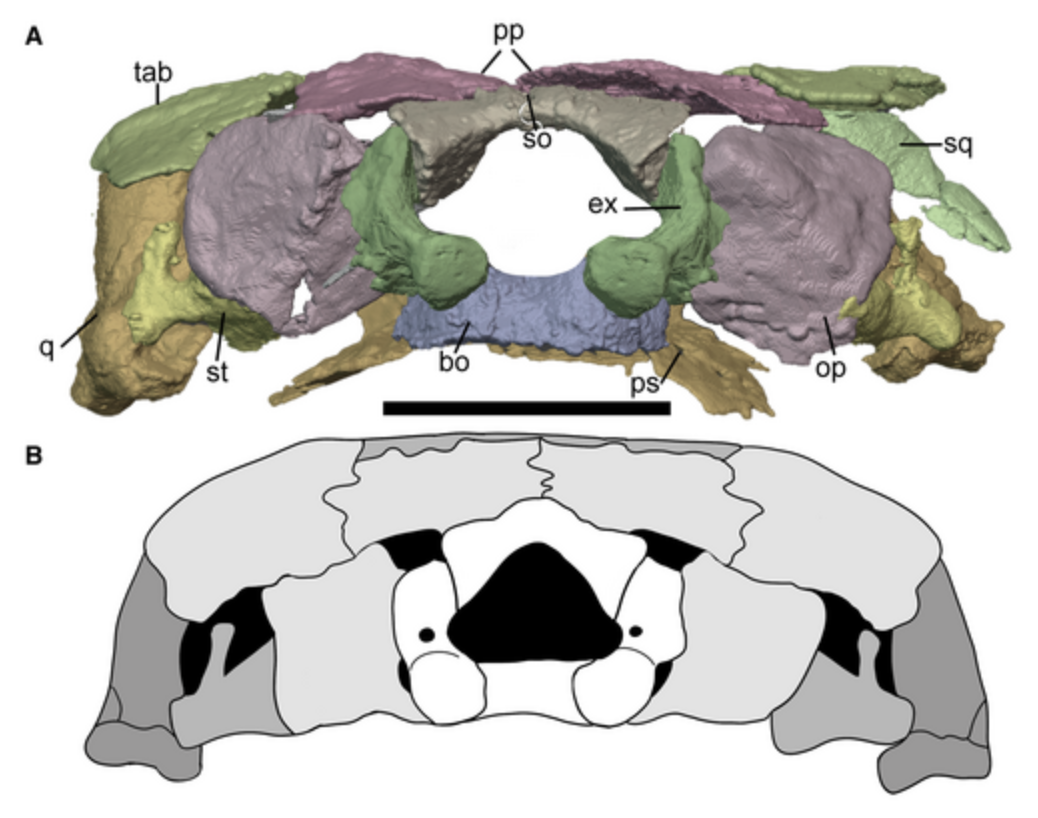
Dorsal views of the braincase of sauropsid Captorhinus (left) and
recumbirostran Rhynchonkos (right) from Szostakiwskyj et al., 2015
showing lateral and median ascending processes (LAP and MAP).A new mess: Captorhinidae
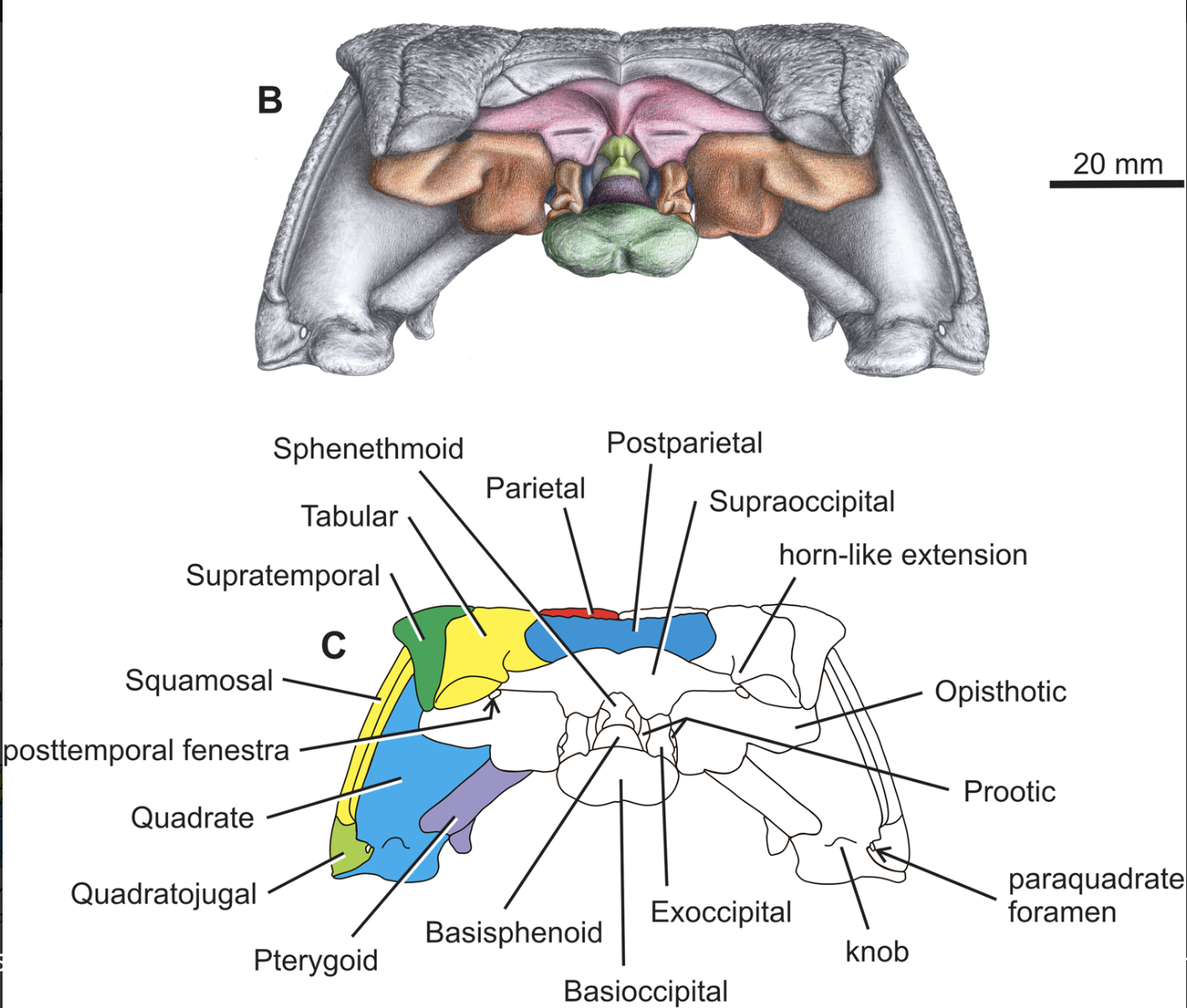
Recumbrostrans like Rhynchonkos were covered above, along with concerns indicated in recent redescriptions that they are actually sauropsid amniotes. Evidence:- Goes beyond the presence of an unambiguous supraoccipital element in the braincase, universal in amniotes.
- Detailed descriptions of three recumbirostrans by Szostakiwskyj et al., 2015 noting that their supraoccipitals resembled those of basal sauropsids in detail, including the presence of lateral and median ascending processes.
- New redescriptions accompanied by phylogenetic analysis supporting the sauropsid hypothesis. (Mann et al., 2022)
- Captorhinidae: (Late Carboniferous - Late Permian) Eureptiles characterized by adaptations to a strong, slow bite with trends toward rounded crushing teeth in multiple tooth-rows. Including Eocaptorhinus (right) but also larger forms such as Labidosaurikos (~1.5 m).
Synapomorphies:
- Complete loss of the ectopterygoid.
- Complete loss of the tabular.
Synapomorphies of Captorhinidae and Amniota:
Limnoscelis, a diadectomorph, skull © Stuart Sumida- Frontals participate in margin of orbit.
- Tabulars are vanishingly small or absent.
- Scapulocoracoid ossifies as scapula, procoracoid (aka "anterior coracoid), and metacoracoid (aka "posterior coracoid") (as seen in Captorhinus). What happens to them?
- All three elements are visible in stem amniotes and persist in basal members of both amniote lineages.
- Synapsida: Traditionally, the procoracoid is thought to diminish among synapsid amniotes, leaving the metacoracoid as a discrete ossification and eventually the "coracoid process" of the scapula in therian mammals.
- Sauropsida: Traditionally the metacoracoid was seen to diminish among basal sauropods, leaving only the procoracoid, usually just called the "coracoid."
 ->
-> 
Before and after:
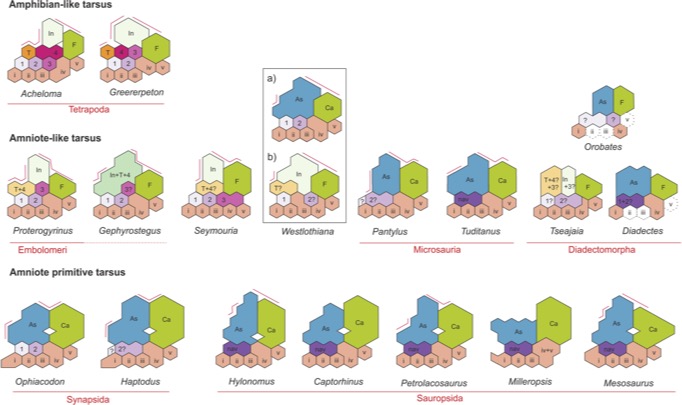
Eryops - a temnospondyl and a generic amniote showing astragalus in aqua - Tibiale, intermedium, and fourth central fuse to form astragalus ("talus" in human anatomical nomenclature.)Various patterns of fusion of the ancestral tetrapod proximal tarsals are seen on the reptiliomorph stem, however among amniotes the pattern is stabilized and characterizes post-hatching life stages in most taxa. (Obviously not snakes.) Similar patterns characterize full adults in some "microsaurs," recumbirostrans, diadectomorphs, and captorhinids.
Points of Interest:
- The oldest fossilized skin of any cotylosaur probably belonged to a captorhinid. Recovered from Early Permian fissure infilling of a cave in Devonian rock, the skin shows non-overlapping epidermal scales - I.e. scales homologous to those of amniotes, not fish or amphibians. (Mooney et al., 2024.)
- The first evidence of caudal autotomy - the ability to detach one's tail to avoid predators - is recorded in captorhinids (leBlanc et al., 2018.) In captorhinids, as in living squamates, this is achieved by the development of pre-exisiting fracture planes that transversely divide the centra of caudal vertebrae like perforations in a sheet of paper.
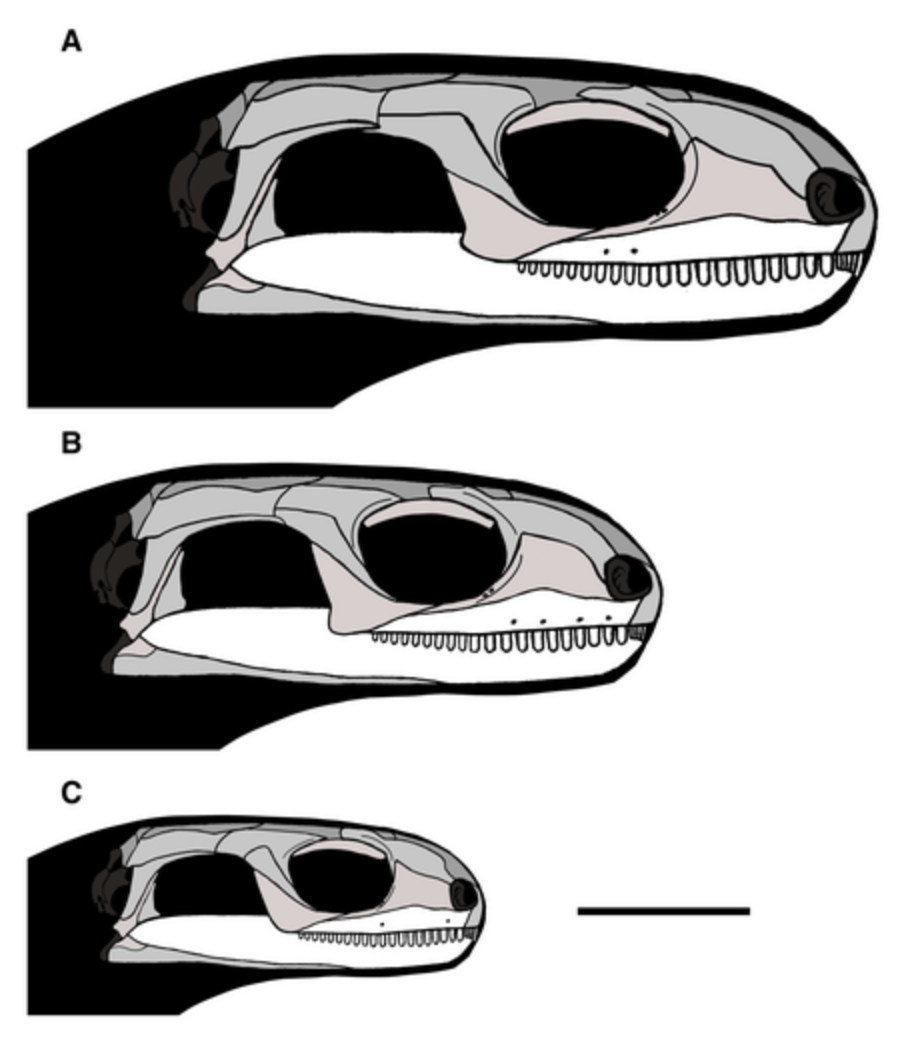
- "Protorothyridids": (Late Carboniferous - Early Permian) Smallish tetrapods with incipient adaptations to a weaker, quick bite. Like captorhinids, considered proper sauropods from the earliest days of cladistics, but now a consensus of new analyses places them on the amniote stem, just outside Amniota, either as sister taxon to Captorhinidae (Simões et al., 2022) or as the sister-taxon to Amniota Jenkins et al., 2025. These include:
- Paleothyris acadiana (right)
- Hylonomus lyelli, from Joggins, NS. Arguably the earliest known amniote, but co-occurs with the fragmentary Protoclepsydrops, which might be the earliest synapsid.
Potential synapomorphies with Amniota:
- Caniniform teeth present
- Rounded occipital condyle (See plesiomorphic condition in Diadectes)
Noteworthy plesiomorphies of Cotylosauria:
- The stapes is robust.
- There is no suggestion of an otic notch.
A parting irony: Before cladistics, the term "Cotylosauria" referred to a waste-basket grade-group taxon of vaguely amniote-grade tetrapods, including captorhinids, protorothyrids, diadectomorphs. The term was resurrected in a phylogenetic sense to encompass Diadectomorpha and Amniota. Now, with the demotion of captorhinids and protorothyrids, the traditional "cotylosaurs" are reunited in a phylogenetic context as an "almost amniote" grade.
And so we come to:
Amniota
The crown group of Reptiliomorpha - last common ancestor of birds and mammals and all descendants. Once, we would have recited a great pile of synapomorphies, however as our understanding of the reptiliomorph stem has improved, all of these have been distributed down the tree. So is Amniota diagnosable? Interestingly, yes.
Archaeothyris - a synapsid and Petrolacosaurus, a sauropsid (probably) by Hugo Karala.Synapomorphies of Amniota:
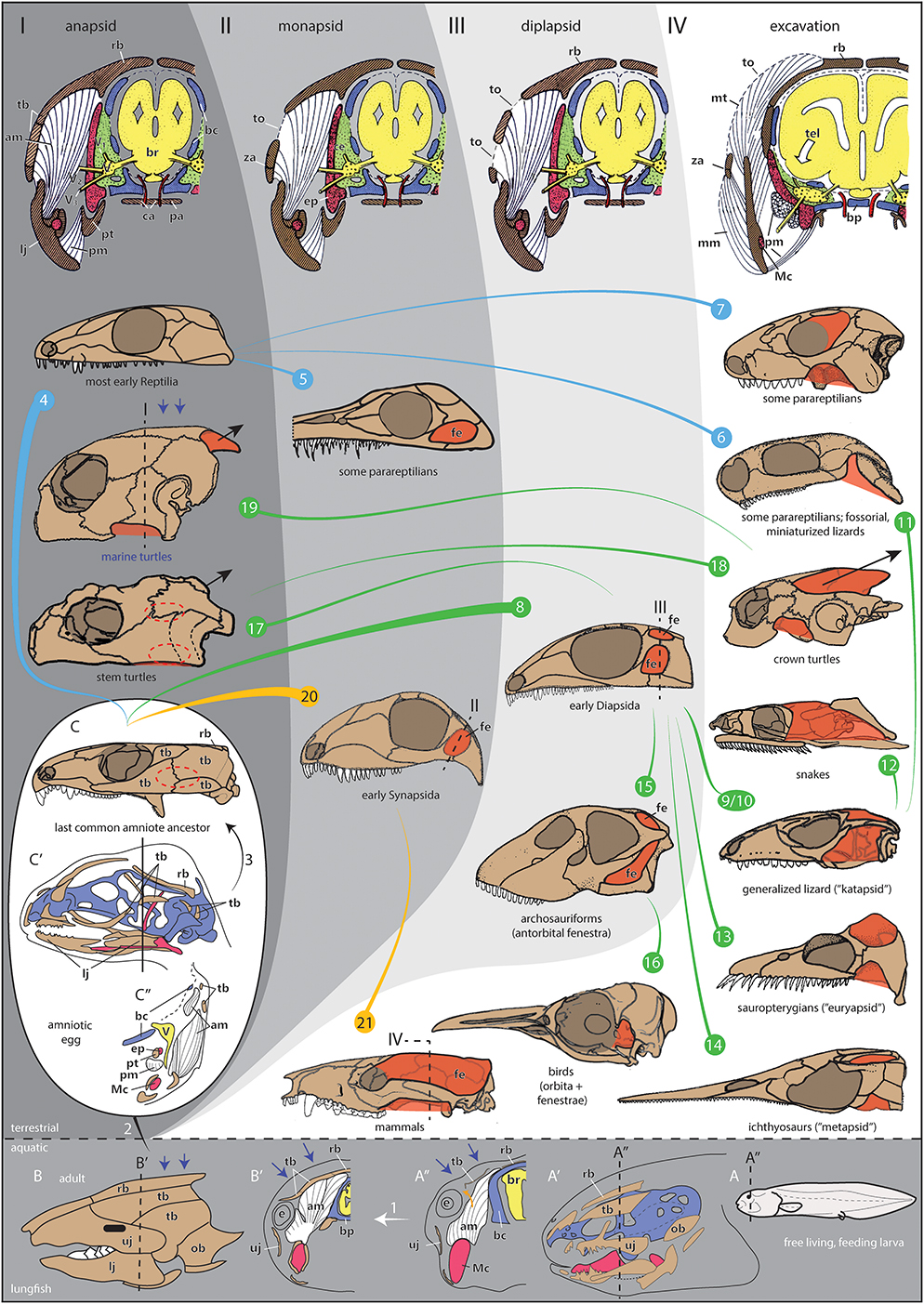
- Infratemporal fenestra. Aka "lower temporal fenestra." This has been considered a synapomorphy of Synapsida since the earliest days of cladistics, however the removal of morphologically anapsid creatures like captorhinids and protorothyridids from Sauropsida to the amniote stem means that the most basal sauropods also possessed it.(Jenkins et al., 2024) But careful! In Synapsida, the infratemporal fenestra is a persistent feature. Among sauropsids, it is often modified by secondary closure or confluence with other openings. A topic for the future.
But first: Cranial fenestration - an amniote trend:Among amniotes, we quickly see the evolution of various patterns of cranial fenestration - the evolution of openings in the skull roof that serve a range of mechanical functions including:
- Allowing the outward bulging of jaw muscles to facilitate a stronger bite.
- Avoiding the waste fo metabolic energy of creating skull-roof dermal bone where it is not needed.
Additional reading:
- Kayla D. Bazzana-Adams, David C. Evans, Joseph J. Bevitt, Robert R. Reisz, 2023. Neurosensory anatomy and function in Seymouria. Journal of Morphology, 284 (5).
- Robert Carroll, 1970. The ancestry of reptiles. Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences. 257 (814): 267Đ308.
- Robert Carroll, 2009. The Rise of Amphibians: 365 Million Years of Evolution. Johns Hopkins University Press, Baltimore. 360 pp.
- Marylène Danto, Florian Witzmann, Johannes Müller, 2012. Redescription and phylogenetic relationships of Solenodonsaurus janenschi Broili, 1924, from the Late Carboniferous of Nýřany, Czech Republic. Fossil Record 15 (2) 2012, 45Đ59.
- Jacques Gauthier, Arnold G Kluge, Timothy Rowe. 1988. Amniote phylogeny and the importance of fossils. Cladistics 4(2):105-209.
- Xavier A. Jenkins, Roger B. J. Benson, David P. Ford, Claire Browning, Vincent Fernandez, Kathleen Dollman, Timothy Gomes, Elizabeth Griffiths, Jonah N. Choiniere, Brandon R. Peecook, 2024 (preprint). Stepwise Assembly Of Crown Reptile Anatomy Clarified By Late Paleozoic Outgroups Of Neodiapsida. Bioarchive doi: https://doi.org/10.1101/2024.11.25.625221.
- Xavier A. Jenkins, Hans-Dieter Sues, Savannah Webb, Zackary Schepis, Brandon R. Peecook, Arjan Mann, 2025. The recumbirostran Hapsidopareion lepton from the early Permian (Cisuralian: Artinskian) of Oklahoma reassessed using HRμCT, and the placement of Recumbirostra on the amniote stem. Papers in Palaeontology 11(1).
- A. R. H. LeBlanc, M. J. MacDougall, Y. Haridy, D. Scott, and R. R. Reisz. 2018. Caudal autotomy as anti-predatory behaviour in Palaeozoic reptiles. Nature, Scientific Reports 8(3328).
- Hillary Maddin and Jason Anderson. 2012. Evolution of the Amphibian Ear with Implications for Lissamphibian Phylogeny: Insight Gained from the Caecilian Inner Ear. Fieldiana Life and Earth Sciences. 5 :59-76.
- Hillary Maddin, Farrish Jenkins, Jason Anderson. 2012. The Braincase of Eocaecilia micropodia PLOS One, December 2012. DOI: 10.1371/journal.pone.0050743.
- Arjan Mann, Jason D Pardo, and Hans-Dieter Sues. 2022. Osteology and phylogenetic position of the diminutive ÔmicrosaurŐ Odonterpeton triangulare from the Pennsylvanian of Linton, Ohio, and major features of recumbirostran phylogeny. Zoological Journal of the Linnean Society 2022;, zlac043,
- David Marjanovic and Michel Laurin. 2019. Phylogeny of Paleozoic limbed vertebrates reassessed through revision and expansion of the largest published relevant data matrix. PeerJ 6:e5565
- Angela Milner. 1988. The relationships and origins of living amphibians. In: M. Benton ed. The phylogeny and classification of tetrapods, volume I: amphibians, reptiles, birds. Clarendon Press. Oxford. 59-102.
- Ethan Mooney, Tea Maho, R. Paul Philp, Joseph Bevitt, Robert Reisz. 2024. Paleozoic cave system preserves oldest-known evidence of amniote skin. Current Biology 34(2).
- Nyakatura, J. A.; Melo, K.; Horvat, T.; Karakasiliotis, K.; Allen, V. R.; Andikfar, A.; Andrada, E.; Arnold, P.; Laustršer, J.; Hutchinson, J. R.; Fischer, M. S.; Ijspeert, A. J. 2019. Reverse-engineering the locomotion of a stem amniote. Nature 565, 351Đ355.
- Jason D. Pardo, Matt Szostakiwskyj, Per E. Ahlberg & Jason S. Anderson, 2017. Hidden morphological diversity among early tetrapods. Nature 546, 642Đ645.
- Ponstein, Jasper; MacDougall, Mark J.; Fröbisch, Jörg, 2024. A comprehensive phylogeny and revised taxonomy of Diadectomorpha with a discussion on the origin of tetrapod herbivory. Royal Society Open Science. 11 (6)
- Laura B. Porro, Elizabeth Martin-Silverstone, and Emily Rayfield, 2024. Descriptive anatomy and three-dimensional reconstruction of the skull of the tetrapod Eoherpeton watsoni Panchen, 1975 from the Carboniferous of Scotland. Earth and Environmental Science Transactions of the Royal Society of Edinburgh. Published online 2024:1-21. doi:10.1017/S175569102300018X
- Tiago Simões, Christian Kamerer, Michael Caldwell, Stephanie Pierce, 2022. Successive climate crises in the deep past drove the early evolution and radiation of reptiles. Science Advances. 8 (33)
- T. R. Smithson, R. L. Carroll, A. L. Panchen and S. M. Andrews, 2017. Westlothiana lizziae from the Visean of East Kirkton, West Lothian, Scotland, and the amniote stem Earth and Environmental Science Transactions of The Royal Society of Edinburgh, 84(3-4)
- Matt Szostakiwskyj, Jason D. Pardo, Jason S. Anderson. 2015. Micro-CT Study of Rhynchonkos stovalli (Lepospondyli, Recumbirostra), with Description of Two New Genera. Plos|One June 10, 2015.
- Matthew K Vickaryous, Brian K Hall. 2006. Homology of the reptilian coracoid and a reappraisal of the evolution and development of the amniote pectoral apparatus. Journal of Anatomy 208(3):263Đ285
- Sebastian Voigt, Gabriela Calabkova, Izabela Ploch, Vojtech Nosek, Wojciech Pawlak, Pawel Raczynski, Frederic Spindler, and Ralf Werneburg. 2024. A diadectid skin impression and its implications for the evolutionary origin of epidermal scales. Biology Letters 20(5)
- Ingmar Werneburg. 2019. Morphofunctional Categories and Ontogenetic Origin of Temporal Skull Openings in Amniotes. Frontiers in Earth Science Sec. PaleontologyVolume 7 - 2019
- Robert Carroll, 1970. The ancestry of reptiles. Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences. 257 (814): 267Đ308.
- Transverse flange of pterygoid. This feature serves as the origin of the medial pterygoideus muscle, jaw adductors whose action is roughly perpendicular to that of the temporalis muscles originating on the braincase and skull roof. Its presence indicates the ability to maintain constant bite force regardless of the position of the jaw - a feature that enhances feeding strategies. But watch out! Porro et al., 2024 identify a transverse process in the pterygoid of the embolomere Eoherpeton, so we may have a poor understanding of this feature's distribution.

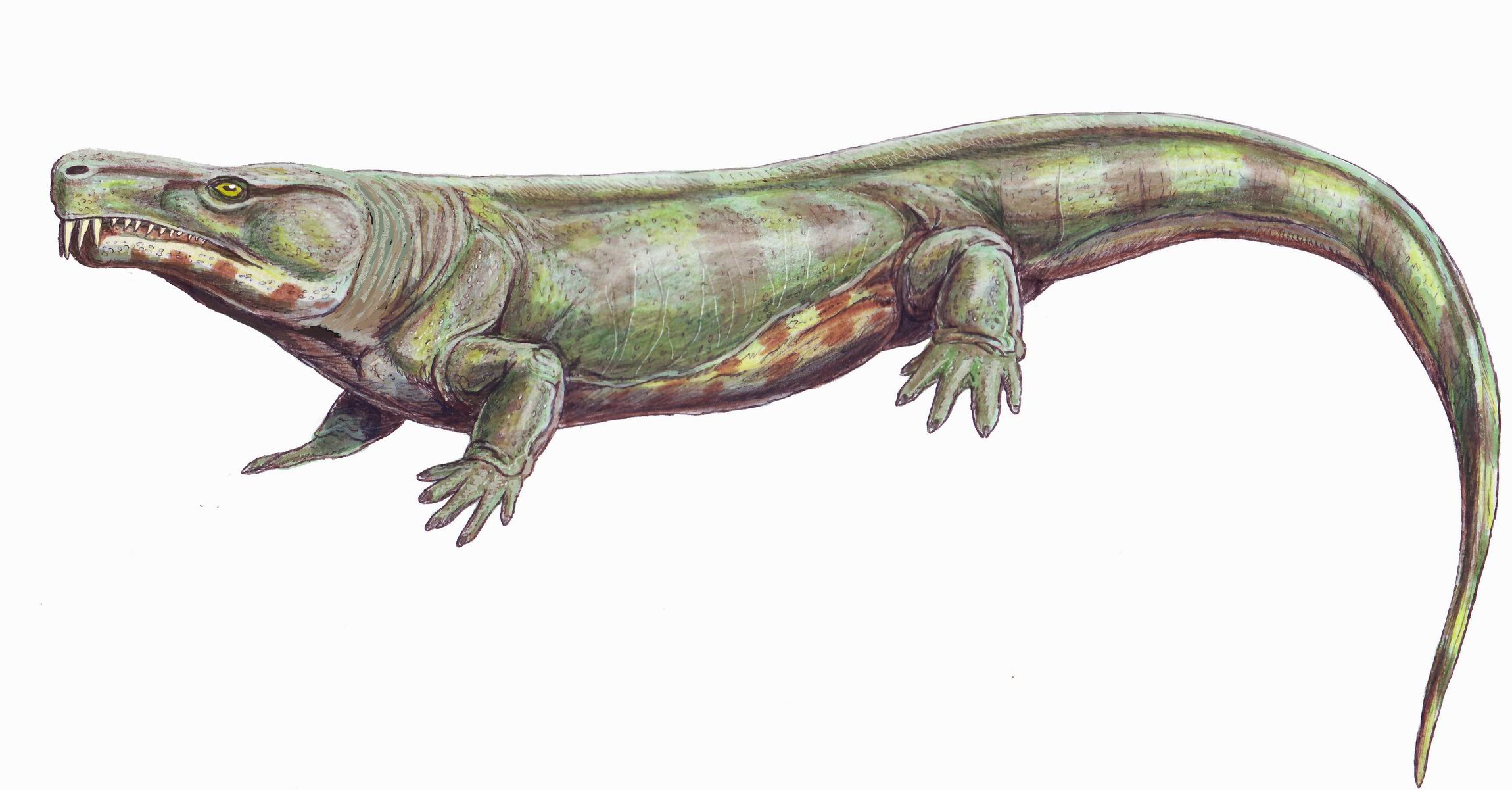
Rhynchonkos stovalli, a microsaur (left) and Paleothyris acadiana, an amniote (right) from Carroll 2009















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}