
Slow Diffusion onto Land
John Merck
Link to Temnospondyli cladogram and phylogram cheat-sheets
Link to Reptiliomorpha cladogram and phylogram cheat-sheets
Elpistostegalia becomes "digitized"


Parmastega has enlarged, dorsally placed orbits, and seems to have hunted with its eyes and skull table protruding from the water. It's nostrils faced downward, but like other elpistostegalians, it could have breathed through its spiracles. It's forelimbs aren't preserved, but its pectoral girdle resembles those of later elpistostegalians with digits. Known only from its skull and girdle elements, Parmastega lacks opercular elements and the dermal pectoral elements dorsal to the anocleithrum. Thus, like Tiktaalik, it had a neck of sorts. Potential synapomorphy with Tetrapoda:
- Paired nasals
When we meet elpistostegalians again in the latest Devonian, O2 concentrations had returned to familiar levels, but air-breathing adaptations had not gone away. Progressing into the Carboniferous - the "Age of Oxygen," the atmosphere became the place for easy access to O2, creating a selective pressure to exploit air-breathing abilities.
We now encounter creatures with very tetrapod-like heads including Ventastega curoni (Ahlberg et al., 2007).
What kind of appendages attached to the pectoral girdle? Unknown. Fortunately in the next case, it is not.

Very well known from relatively complete skeletons. The first and phylogenetically most basal vertebrate definitely to possess fingers and toes! Synapomorphies with Tetrapoda include:
- Nasals nearly touch on mid line (although a narrow internasal fontanelle remains.)
- Ethmosphenoid and otico-occipital components of neurocranium are fused. Intracranial hinge obliterated.
- Pharyngeobranchials lost.
- Fingers and toes present
Acanthostega's neurocranium at last begins to resemble that of a tetrapod. Note:

- Distinct basipterygoid processes compared to what we have seen previously.
- Ventrally, the lateral cranial fissure manifests as a large vestibular fontanelle (fen vest right).
But was it a fish? Consider the transformations listed at the beginning of the lecture:

- Breathing
- Osteology of the pectoral girdle tells us that an open operculum and functional gills were present in adult. (Although they were reduced slightly in size by the loss of phanyngeobranchials. (Schoch and Witzmann, 2011.)) Specifically:
- Ceratobranchials were grooved posteriorly to accommodate branchial arteries
- The cleithrum had a clear postbranchial lamina.
- The tabular enclosed a large spiracular opening.
- Osteology of the pectoral girdle tells us that an open operculum and functional gills were present in adult. (Although they were reduced slightly in size by the loss of phanyngeobranchials. (Schoch and Witzmann, 2011.)) Specifically:
- Mechanical support
- Flexibility of forelimb
- As modeled digitally by Molnar et al., 2021,
- strength of humerus retraction is intermediate between that of lungfish and Eusthenopteron and of living tetrapods, suggesting some ability to generate force against a substrate.
- The elbow joint is highly mobile
- But the humerus can't be rotated to anything like the extent of living tetrapods.
- Slightly flexible elbow and knee.
- Fin rays lost from all but the caudal fin
- Fingers (eight per limb) and toes (seven per limb) present.
- accessory articulations between neural arches, but no zygapophyses to speak of.
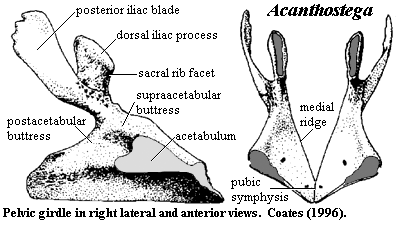
- Pelvis is small but has a ligamentous connection with a sacral rib.
- But note: It is not merely the presence of digits and mobile joints that makes a terrestrialized limb. The number and mass of muscles and nerves to overcome them has to increase significantly. Hirasawa et al., 2021 investigated the development of the forelimb in the Australian lungfish Neoceratodus, finding that a limiting factor to forelimb enlargement was that amount of mesenchyme able to migrate into the limb to form its peripheral nervous system. The early ossification of the cleithrum seems to form a barrier to mesenchyme migration, limiting forelimb mass. Consequently, the elaboration of the forelimb was coupled evolutionarily with reduction of the dermal cleithrum.
- Feeding:
- Skull and pectoral girdle detached.
- First vertebra is enlarged, but otherwise no specialization of the vertebral column, such as zygapophyses.
- Skull and pectoral girdle detached.
- Senses:
- Extensive lateral line system
- Unlike in other gnathostomes we have considered so far, the foot-plate of the hyomandibula rests in the vestibular fontanelle, rather than articulating to a solid facet on the neurocranium. In better-known tetrapods, this is how sound is transmitted to the otic capsule. When it takes on this function, the opening is known as the fenestra vestibuli or fenestra ovalis.
- Vision out of the water necessitates blinking. Land vertebrates have eyelids or nictitating membrane for blinking, however non-amniotes, including mudskippers and amphibians blink in whole or in part by retracting the eyeball into the eye socket using the retractor bulbs muscle, whose osteological correlates are present in Acanthostega. (Aiello et al., 2022.)

The overall impression is of a fully aquatic animal with rudimentary ability to support some body weight on fore and hindlimbs and with the beginnings of the ability to use its jaw and hyoid arch to pick up sound from the ground. Maybe it came onto land occasionally to feed, disperse to new habitats, or thermoregulate, but it was primarily an aquatic animal. What did it use its fingers and toes for? Maybe not much at all. As with the origin of lungs in Osteichthyes, Byrne et al., 2020 speculate that the evolution of digits might be a response to high tide ranges and the hazards of becoming trapped in tide pools.

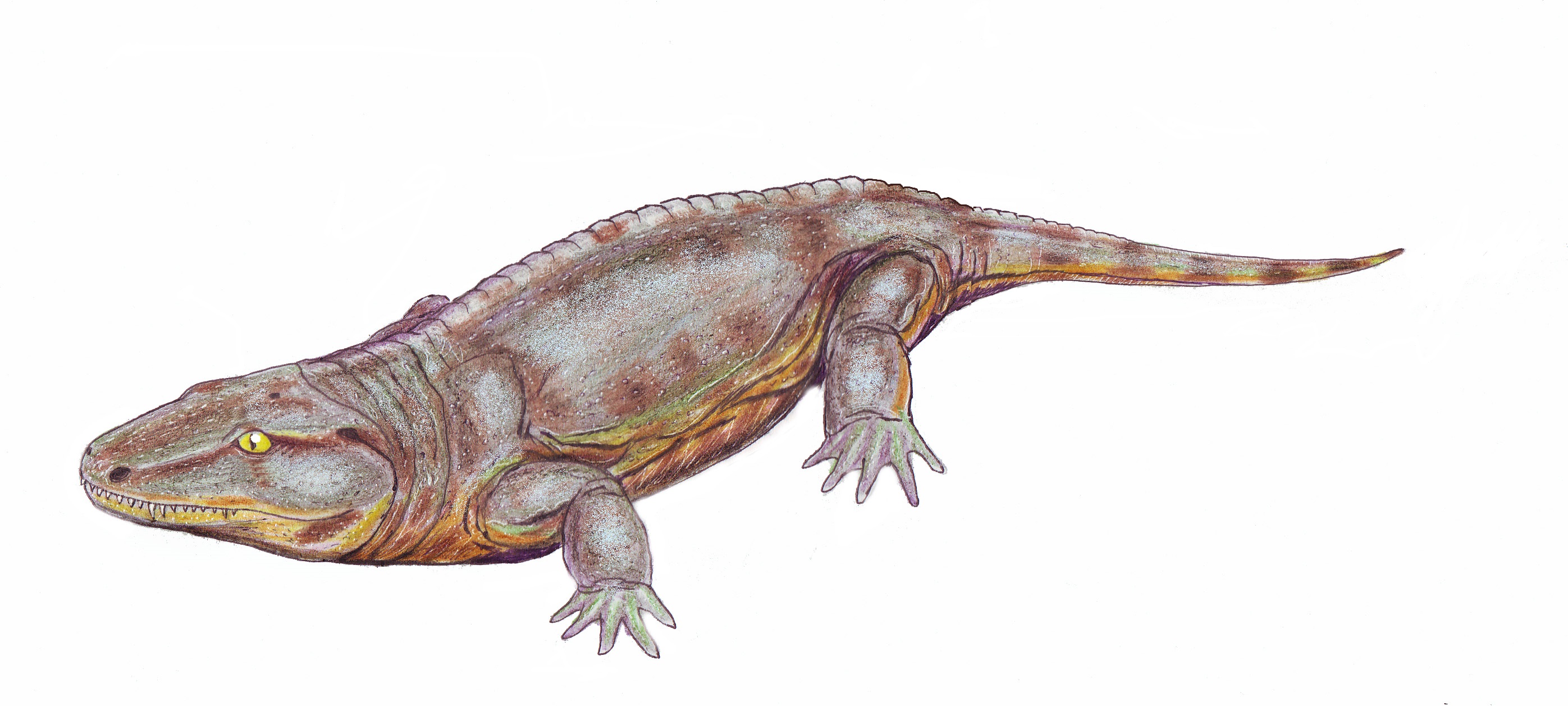
Ichthyostega stensioi
Very well known (even in popular culture of previous generations: See Album cover of Yes' "Tales of Topographic Oceans" by Roger Dean) from relatively complete skeletons, but redescribed by Ahlberg et al., 2005.
- Breathing
- Small detached opercular element and grooved ceratobranchials remained (Clack et al., 2003), indicating that it breathed using internal gills.
- Mechanical support
- Ribs are long broad and overlapping, limiting ability to curve spine laterally either in water or on land. Instead, the regions of the vertebral column are specialized, sort of like in a mammal. Implication is that spine was flexed up and down.
- Zygapophyses present. (But note the reversed positions of intercentra and pleurocentra!)
- Pelvis robust in comparison to all aquatic vertebrates, with direct articulation with sacral rib.
- Ribs are long broad and overlapping, limiting ability to curve spine laterally either in water or on land. Instead, the regions of the vertebral column are specialized, sort of like in a mammal. Implication is that spine was flexed up and down.
- Seven toes present, as in Acanthostega. The earliest creatures with fingers and toes had the developmental ability to generate them, but had not yet evolved the ability to stop making them when they had enough.
- Flexion of elbow and knee joints limited. The limbs seem to have functioned primarily as paddles.
- Feeding:
- Skull and pectoral girdle completely detached. Dermal pectoral series dorsal to the cleithrum completely lost.
- But there is still a pit for the notochord in the basioccipital condyle, limiting head's ability to flex.
- Senses:
- Lateral line system remains present
- The hyomandibula is weird in this animal, being very broad, flat and thin. Apparently is had some unique function that is not seen in living vertebrates. Clack et al., 2003 speculates its flat surface rested against an air-filled chamber in the spiracular passage, picking up airborn vibrations and transfering them to the otic capsule via the fenestra vestibuli. In that way, it functioned analogously to an eardrum in an early attempt at an impedance-matching ear. In that case, how likely is it that Ichthyostega also had an ear-drum?
- Lateral line system remains present


- Reduced operculum
- strengthened vertebral column
- reduced ability to flex body laterally

Tulerpeton curtum scale bar = 1 cm.
- Absence of evidence for an open operculum.
- A six-fingered hand
Elegant Hypothesis - Brutish Facts: We have constructed an attractive and coherent picture of the steps in the acquisition of tetrapod-like attributes:
- Late Devonian oxygen concentration crisis stimulates the evolution of enhanced air-breathing in elpistostegalians.
- Latest Devonian acquisition of limbs with fingers and toes combines with existing air-breathing habits to yield a vertebrate capable of venturing onto land.
-
Middle Devonian trackways: The footprints of quadrupedal creatures with fingers and toes are known from several localities around the world. Although the geologic age of many are poorly constrained, some (see Niedzwiedzki et al., 2010) are plainly Middle Devonian - i.e. 18 my older than Acanthostega. WTF? Apparently Elpistostagalia encompasses some substantial ghost lineages.

Romer's Gap - a geologic enigma: Acanthostega and Ichthyostega lived roughly 365 mya. The earliest known members of the lineages leading to Lissamphibia and Amniota are from roughly 335 mya. For some reason, the first 30 my of the Carboniferous were a dark age in which fewer were preserved as fossils or paleontologists haven't found the correct rocks. (Note: a similar contemporaneous gap exists for fossil insects.) Thus, we are in the dark about how quickly vertebrates actually used their hands and feet to get out of the water. Even after Romer's Gap ended, most "land vertebrates" remained aquatic with weak limbs and prominent lateral line systems.
Chastened, we consider the Carboniferous: Remember, the latest Devonian "digitized" elpistostegalians weren't, as far as we can tell, really using their digits for much. From these beginnings, creatures had three general options:
- Reduce or eliminate limbs and digits
- Continue to not specialize (ignore) them
- Develop them for new functions

Oestocephalus amphiuminus
(Early Carboniferous - Early Permian) Superficially snake like, elongate, and limbless (Although Phlegethonia retains a cleithrum in the pectoral region (Anderson, 2002). Long regarded as members of crown Tetrapod, Pardo et al., 2017 found to be early stem tetrapods (ironically) based on ancestral features including:
- Presence of a notochord recess in place of basioccipital condyle.
- Open lateral cranial fissure
- Persistent hypophyseal duct opening in palate.
Skulls are significantly modified, with orbits shifted far anteriorly and jaw articulation shifted posteriorly resulting in the great elongation of the temporal region. What's particularly strange is the reduction and fragmentation of the dermal bones of the temporal region. To compensate for the loss of mechanical strength that this implies, the palatoquadrate actually ossifies as a single unit.

Aistopod vertebrae are holocentrous, however Lethiscus, from the Early Carboniferous, was found by Anderson et al., 2003 to retain distinct intercentra and pleurocentra. It seems to be the most primitive aistopod. Alas, being so highly derived, the paleobiology of aistopods is difficult to interpret. Because descriptions of branchial arches and pectoral girdle (such as it is) are lacking, we can't even assess whether they were primarily air breathing.
Whatcheeridae: (Early - early Late Carboniferous) Clack, 2002 described Pederpes finneyae of Britain, from the middle of Romer's Gap at 350 mya. Although it retains a tiny sixth finger, has a robust lateral line system, a heavy hyomandibula/stapes, and retains a preopercular; it also has robust limbs and a skull that is compressed from side-to-side like early land vertebrates. It and close relative Whatcheeria comprise the Whatcheeriidae (Latest Devonian - Early Carboniferous). We don't know how much time they spent on land, but they seem, at least, to have been able to walk - a new development fifteen million years after the evolution of hands and feet. Otoo et al., 2021 note that in comparison to other basal tetrapods, whatcheeriids have extremely large robust limbs, but that they present plausible adaptations to aquatic life. On the other hand, Molnar et al., 2021 note that in their simulations, Pederpes is only slightly advanced over Acanthostega.

Early Carboniferous Landmarks: Emerging from Romer's Gap, we pick up several lineages. Their synapomorphies:
- The preopercular - the last vestige of the dermal opercular skeleton - is lost.
- Opening of the hypophyseal duct in parasphenoid lost.
- Notochordal pit in basioccipital is lost.
Colosteidae: (Carboniferous - Early Permian) Including the familiar Greererpeton. All are elongate with small limbs whose joints are formed in cartilage. They retain extensive lateral line systems, and some have gill-rakers on their ceratobranchials, indicating an open operculum and suspension feeding as an option (Schoch and Witzmann, 2011). Distinctly fresh-water aquatic. Distinctive features:

- Spiracular notch is lost. (Spiracle likewise?)
- Prefrontal extends to margin of naris
- Premaxillary fang fits in notch in dentary
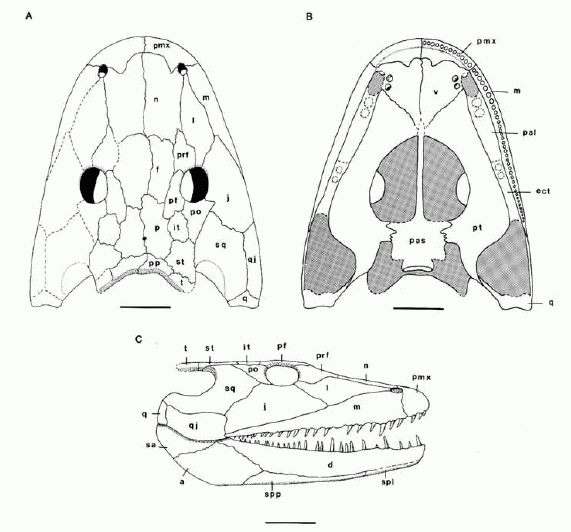
Crassygyrinus scoticus: (Early Carboniferous) Very strange but deeply rooted in popular culture:

- The body is elongate with a deep tail.
- Hindlimbs are small, forelimbs positively puny.
- Sacral articulation lost
- Vertebral column is poorly ossified, with poorly developed zygapophyses.
- As reconstructed by Panchen 1973 and Clack, 1998 the head was strongly laterally compressed such that the eyes definitely looked to the side. Redescription by Porro et al. 2023 (right) indicates that this is exaggerated and that the head was actually flatter.
- Models of jaw joint indicates a gape of up to 60 degrees.
- A distinct spiracular notch
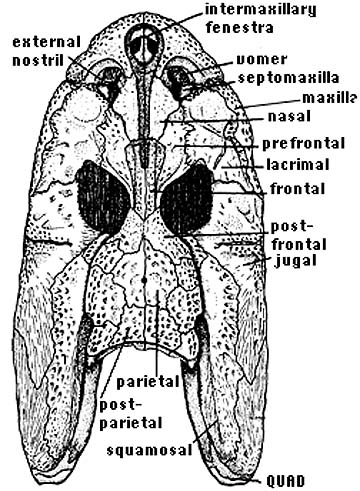
- A midline trough on the snout, the intermaxillary fenestra. Is this the internasal fontanelle we saw in Ventastega?
Baphetidae:(Carboniferous) A morphologically disparate but distinct clade of aquatic predators. Known since the 19th century from skulls but hardly any postcranial material. Plesiomorphic in retention of full lateral line system and distinct spiracular notch.

Synapomorphies:
- Solid palate with closed interpterygoid vacuities and coossified basipterygoid articulations.
- Key-hole shaped orbit with embayment of the lacrimals and prefrontals.
- a gland?
- a fenestra to accomodate bulging muscles?
Baphetid diversity: encompasses several ecomorphs.
- Eucritta melanolimnetes (Early Carboniferous) Might be the most basal baphetid. Surprisingly small (~25 cm) and with only a hint of the key-hole orbit. (Psst! Also recovered as sister taxon to Tetrapoda and stem amphibian.)
- Megalocephalus with a superficially crocodile-like skull up to 35 cm long. The largest of a range of fish-eating aquatic predators. In it, the basipterygoid articulation becomes a solid suture uniting palatoquadrate and neurocranium.
- Spathicephalus: Evidence of hidden diversity. Shovel-headed with exaggerated key-hole orbits and unprecedented dentition of many chisel-like teeth that don't appear to undergo replacement, and no palatal teeth at all. Could this be a ram-suspension feeder (like a baleen whale)? In all its weirdness, on tends to overlook that it, too, fuses the basipterygoid articulation.
Phylogenetic analyses place baphetids either below the base of crown Tetrapoda or as basal members of the Lissamphibian stem. A potential synapomorphy of baphetids and the lissamphibian line: A slender hyomandibula suspended in the spiracular chamber - except now we can start saying "A slender stapes suspend in the middle ear."
Anthracosauria: (Carboniferous - Triassic) Aquatic to mostly terrestrial vertebrates of the late Paleozoic. Although this groups membership varies with each new publication, it seems monophyletic.

Identifying characters:
- The tabular contacts the parietal and develops elongate "horns" at the posterolateral corner.
- The cheek lies adjacent to the skull roof along a kinetic line.
- There is no clear spiracular notch, and the stapes is heavy. Sensory canals present in the skulls of many.
- Two - three fenestrae perforate the medial surface of the jaw.
- Pleurocentra larger than intercentra . Both assume a horseshoe or disk shape.

Relationships: Until recently seen as the basal members of Reptiliomorpha. Some characteristics, including the tabular parietal contact and the large pleurocentra are shared with other reptiliomorphs, including amniotes. However Clack, 2011, suggested, based on the persistence of plesiomorphies like caudal fin rays (Clack, 2012), that hey might belong on the Tetropod stem. This was confirmed by Pardo, 2024 in his redescription of the embolomere Archeria.
Final word:
Stem tetrapods are paradoxical. All show adaptations indicating increased reliance on air-breathing, the hearing of airborne sound, and some may have routinely walked on land: however:
- Acanthostega, Greererpeton, Whatcheeria, and Megalophalus, all display osteological correlates to the presence of internal gills (Schoch and Witzmann, 2011).
- Modeled limb dynamics indicate that they lacked the land locomotion abilities of living tetrapods. (Molnar et al., 2021)
- Studies of anisotropies in limb bone cross-sections reveal that limb bones of creatures of Acanthostega, Pederpes, and embolomere grade resemble those of living aquatic tetrapods. (Lennie et al., 2021)

Tetrapoda: Opinions vary as to where this name should be stuck to the tree:
- Clack, 2012 feels that excluding creatures with hands and feet from a group whose name means "four feet" is perverse. For her, Tetrapoda has is a classic "apomorphy-based definition."
- Coates et al., 2008 favor restricting the name to the "crown group" - i.e. the node-based group defined as the last common ancestor of living members of the group.
Synapomorphies:
- First two cervical vertebrae specialized as the atlas and axis. These facilitate movement of the head on the neck.
- Presence of a distinct larval stage with three pairs of external gills: Among stem tetrapods like colosteids, open gill slits and fish-like branchial arches are present throughout life, and there is no clear transition between larval and adult forms. Beginning with Tetrapoda, we see specialized aquatic larvae that transition (quickly or slowly depending on the taxon) into adults that may or may not lose their internal gills. Some clades contain neotenic members in which the larvae reach sexual maturity without metamorphosing. (Schoch and Witzmann, 2024.)
- Not a synapomorphy, exactly, but somewhere before Tetrapoda, the number of digits on manus and pes had stabilized at five.
Phylogeny: Tetrapoda consists of two major lineages with living members:
- Amphibia: (total-group) All organisms more closely related to frogs, salamanders, and caecilians than to Amniota.
NOTE: This use of the term "Amphibia" excludes creatures like seymouriamorphs and lepospondyls, even though they might have ecologically resembled modern amphibians.
- Lissmphibia: (Crown-group) The last common ancestor of living amphibians (frogs, salamanders, and caecilians) and all its descendants.
- Reptiliomorpha: (total-group) All organisms more closely related to Amniota than to Lissamphibia.
NOTE: Some creatures in the reptiliomorph total-group, might have ecologically resembled amphibians despite their evolutionary propinquity to amniotes.
- Amniota (Crown-group) The most recent common ancestor of mammals and birds and all of its descendants.
The Amphibian Stem:
Temnospondyli: (Carboniferous - Cretaceous/Quaternary (?)) The name originally coined for basal tetrapods with rhachitomous vertebrae, but has become one of the better-supported monophyletic groups. Fabulously diverse and speciose. Small to large and aquatic to mostly terrestrial. (Morphometric analysis of vertebrae by Carter et al., 2021, suggest that ancestrally, adult temnospondyls were terrestrial but experienced repeated reversions to semi- or fully aquatic ecologies.) Temnospondyls were a primary component of the late Carboniferous and Permian land biota, and experienced a Triassic radiation of aquatic forms. Morphologically they varied from unspecialized to rather stout, short-tailed forms. Very few evolved the eel-like shape so common in embolomeres.

Synapomorphies:
- Four-fingered manus. Note: What's lost is the oddball digit V, not derived from the digital arch. (Pes retains five toes.)
- The humerus has a distinct shaft at mid-length.
- Slender rod-shaped stapes. (Since most also have a large notch at the rear of the skull, most view as evidence for impedance-matching ear.)
- Wide interpterygoid vacuities (link to Capetus scale = 5 cm.) in the palate. In life, the skin covering these was covered by a shagreen of tiny denticles.
- Exoccipitals sutured to postparietals and form paired occipital condyles.
Temnospondyl Diversity: The problems and competing hypotheses of temnospondyl phylogeny are beyond our scope. What follows is a review of the major groups recovered by Schoch, 2013 and Schoch, 2018.
Edopoidea: (Carboniferous - Permian) A basal and plesiomorphic group of large predators, ranging from the more terrestrial Capetus to the aquatic Nigerpeton.

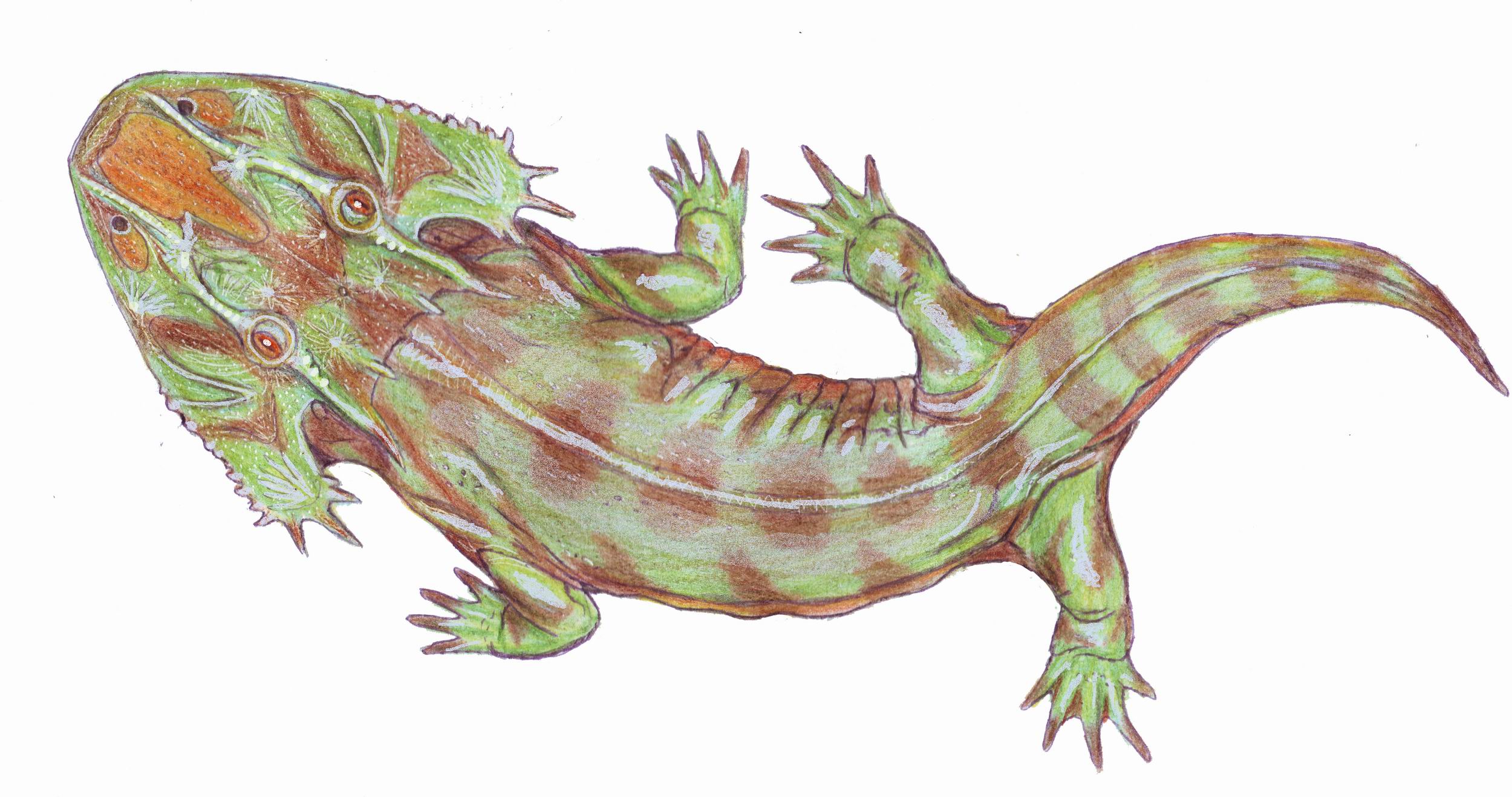
"Dendrerpetontidae": (Carboniferous) A basal and plesiomorphic group of small predators of uncertain monophyly. Interesting because it consists of:

- Dendrerpeton (Late Carboniferous - right). A small form known from well-preserved specimens in Joggins Nova Scotia tree trunk casts. Its skull displays a deep otic notch and wide interpterygoid vacuities. Judging from its lack of sensory canals and preservation in a terrestrial environment, its seems to have been primarily terrestrial as an adult. Its pectoral girdle conveys a good impression of the ancestral state for Tetrapoda.
- Balanerpeton: (Early Carboniferous) The oldest known temnospondyl, also from the lacustrine environment of East Kirkton, Scotland. These, too, seem to be terrestrial animals whose bodies washed into a lake bed.

Capetus - an edopoid (left) and Doleserpeton, a member of Rhachitomi (right)
- Loss of the intertemporal.
- Trend toward enlarged interpterygoid vacuities. (Note: In Doleserpeton (right) the pterygoids no longer meet anteriorly.)
Dvinosauria: (Late Carboniferous - Early Triassic) First representatives of a recurring theme in temnospondyl evolution - paedomorphic adults that retain larval characteristics including gills as an adult. Speciose and common during the Permian.

Zatracheidae: (Late Carboniferous - Permian) Aquatic temnospondyls with flattened spiny armored skulls. An internarial fenestra stimulates idle speculation about its purpose. Link to reconstruction of Zatrachys.

Dissorophoidea: (Late Carboniferous - Quaternary) A diverse group of small temnospondyls including:

- Micromelerpetontidae: (Carboniferous - Permian) Small aquatic forms with healthy sensory canals.
- Olsoniformes: (Carboniferous - Permian) Stout terrestrial forms with dermal armor and deeply incised otic notches (right). This includes the sail-backed temnospondyl Platyhystrix.
- Amphibamiformes: (Carboniferous - Permian) Small forms with large orbits and interpterygoid vacuities. In some ways convergent on amniotes:
- Sensory canals lost.
- Humerus with a long shaft.
- Gastrocentrous vertebrae: with large pleurocentra, often coossified with neural arches, and small crescent-shaped intercentra.

Eryopiformes: (Permian - Cretaceous) The large Mesozoic radiation of aquatic temnospondyls and their basal Permian relatives.
Synapomorphies:
- Otic notch is restricted to the dorsal part of the cheek.
- Prefrontal and jugal in contact in margin of the orbit.
Diversity:
- Eryopidae: (Carboniferous - Permian) The basal group contains Eryops, beloved of illustrators.
- Archegosauridae: (Permian) Specialized long-snouted predators that seem to have hunted in fresh water but ventured onto land frequently in the manner of crocodilians.
- Stereospondyli: (Permian - Cretaceous) Aquatic fresh-water and marine predators that experienced a significant radiation in the Triassic.
Stereospondyl evolutionary trends:- Reduction of the pleurocentra. This reaches its logical conclusion in some members with proper stereospondylous vertebrae.
- Flattening of the head.
- Further enlargement of the interpterygoid vacuity.
- Basipterygoid articulation (formerly a mobile joint) becomes an immobile suture.
- Occipital condyles project far posteriorly.
- Elongate transverse processes and neural spines.
- Elimination of distinct larval and adult life stages. Juveniles look like little adults and both retain internal gills. (Schoch and Witzmann, 2024.)
Stereospondyl diversity:
- Paraphyletic grade of basal members including Rhineceps nyasaensis (right) and Koolasuchus cleelandi, from the Cretaceous of Australia, the youngest known stereospondyl. (Maybe.)
- Trematosauria: (Triassic - Jurassic) Large forms with palate strongly integrated with neurocranium by broader basipterygoid sutures and suturing of pterygoid to exoccipitals. (Link to Thoosuchus yakovkevi.) Spectacular examples:
- Trematosauridae: (Triassic) Long-snouted marine and fresh-water forms. Link to skull of Trematosaurus.
- Metoposauridae: (Triassic) Exceptionally flat-headed lurkers in Triassic fresh waters. (Link to Buettneria perfecta skull and skeletal mount.
- Plagiosauridae: (Triassic - Jurassic) Short-faced paedomorphic forms, yet still large. (Link to skull of Gerrothorax.)
- Capitosauria: (Triassic - Earliest Jurassic) Including the largest heads of any temnospondyl, belonging to the likes of Mastodonsaurus. (Link to skull of Mastodonsaurus giganteus.)

Breathing in Amphibia: Recall that even in stem-tetrapods who relied on lungs to obtain oxygen, Witzmann, 2015 notes that the eliminate CO2 through the gills because this is more efficient in an aquatic environment. But gills impose limitations because they must be kept moist. Tetrapods have found two ways around this problem:
- Lissamphibians exchange gasses including CO2 through the skin. Easy enough in aquatic or moist terrestrial environments.
- Amniotes suck it up and exchange all gasses through the lungs. Presumably the advantages of terrestrial life outweigh the cost of less efficient CO2 elimination.
- Dissorophoids
- Dendrerpetontids
In contrast, adult gills were lost very early on the reptiliomorph side, whose members relied upon lungs for all gas exchange.
Lissamphibia (?) - the living amphibians
Traditionally regarded as monophyletic, Lissamphibia contains all three groups of living amphibians and one fossil group:
- Anura: (Early Triassic - Quaternary) Frogs
- Caudata: (Late Triassic - Quaternary) Salamanders
- Gymnophiona: (Early Jurassic - Quaternary) Caecilians
- Albanerpetontodae: (Jurassic - Neogene)
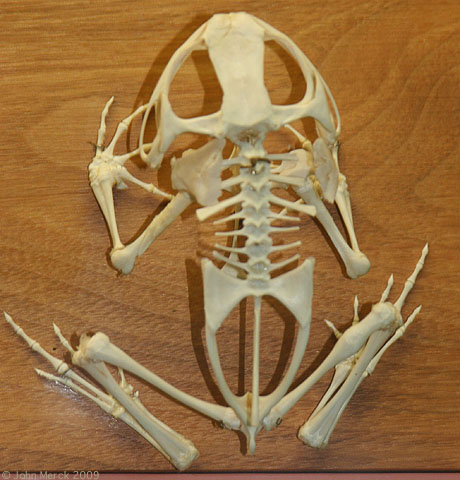
Megophrys montana
Anura:
(Early Triassic - Quaternary) Frogs are very highly derived for specialized forms of locomotion, hearing, and prey-capture. Although most hang out near water, only a few actually feed in the water, and their adaptations are not as useful there. Among their idiosyncrasies:-
Postcranial:
- No more than eight presacral vertebrae.
- Caudal vertebrae ossify as a urostyle - a single long rod.
- The ilium is very long, articulating with the sacral rib anteriorly and the femur posteriorly. Together, these two synapomorphies enable the frog to rotate its torso around its hips, extending the back while jumping.
- Tibia and fibula are fused into a single unit.
- The tibiale and fibulare (proximal tarsals) become elongate.
- Scales completely lost, facilitating cutaneous breathing.
- Frontals and parietals fused into paired frontoparietals.
- Extremely large otic notch accommodates tympanum.
- Jaw muscles reorganized to accommodate large middle-ear cavity.
- Extremely wide interpterygoid vacuities.
- Lower jaw without teeth.
- Hyoid skeleton specialized to allow prey capture by protrusion of the tongue.
- The tadpole - a morphologically and ecologically distinct larval stage. Tadpole fossils known from Middle Jurassic stem anurans demonstrate that this biphasic life-history is fundamental to anurans (Chuliver et al., 2024).
Cranial:

Developmental:
Most fossil frogs share these features (although some have more dorsal vertebrae). Our earliest glimpse is from Triadobatrachus from the Early Triassic. Although plesiomorphic in many ways, and probably unable to jump, it shows the initial stages of many froggie adaptations of the cranium and postcranium.


Caudata:
(Late Triassic - Quaternary) Salamanders. Generally less specialized than other lissamphibians and frequently adapted for life in the water. Although their water-breathing larvae are less specialized, we often see paedomorphic, permanently aquatic salamanders, and many of their anatomical specializations seem like adaptations to aquatic life. As in frogs, scales are completely lost, facilitating cutaneous breathing.Cranial synapomorphies:
- Otic notch and tympanum lost - impedance-matching ear reduced.
- Maxilla does not connect posteriorly with the cheek, creating a gap in the skull margin.
- In basal forms, the squamosal hinges to the neurocranium and can swing laterally, facilitating expansion of the oral cavity and improving aquatic suction-feeding.
Q: Where else have wee seen this?
Postcranial synapomorphies:
- Distal carpals (and tarsals) 1 and 2 fused into a basale commune. This is only the beginning.
- Alone among tetrapods, the chondrification sequence of the digital arch is reversed, with digits I and II forming first, and digit V (!) last. Note that in salamanders, digit V forms as part of the digital arch! (Fröbisch and Shubin, 2011.)
- Chunerpeton (Late Jurassic - China). Regarded as a stem salamander by Rong et al., 2020.
- Karaurus. (Late Jurassic - China)
- Marmorerpeton (Late Jurassic - Scotland) Closely related to Chunerpeton according to Jones et al., 2022..
- Triassurus (Late Triassic - Uzbekistan) A poorly preserved juvenile that, nevertheless, displays the characteristic cheek-gap.

Gymnophiona:
(Late Triassic - Quaternary) Caecilians. As weirdly derived as frogs but in the opposite direction - as limbless burrowers (although some are secondarily aquatic.) Caecilians retain small scales and do not breathe cutaneously.Morphology:
- The skull, although missing many bones, is compact and strong, for use as a digging impliment.
- The eye sockets are greatly reduced, and the eye is often covered by a layer of skin.
- A unique sense organ, the tentacle, protrudes slightly from below the eye. The tentacle can be extended slightly. (In some, the eye and tentacle are consolidated such that the eye can be made to protrude like that of a surprised Loony Toon character.
- Reproduction: Fertilization is internal. (Males have intromittant organ called the phallodium, whose homology to the amniote penis is unknown.) Caecilians can be oviparous (laying large unshelled eggs in moist ground on land) or ovoviviparous.
- Apparently, caecilians are capable of secreting oral venom that can be transmitted into their prey (Mailho-Fontana et al., 2020.)
Features of the skull:

- Otic notch and impedance-matching ear lost.
- Endochondral elements of the neurocranium fused into an os basale
- Bones of the jaw fuse into:
- Pseudodentary anteriorly
- Pseudoanguar posteriorly
- Behind the jaw articulation, a spectacular retroarticular process provides leverage for jaw-closing muscles that originate on the branchial skeleton.
Eocaecilia: (Early Jurassic) Only one fossil caecilian suggests their ancestral form. Noteworthy for the retention of limbs, however many derived caecilian adaptations are clearly visible. Its skull is plesiomorphic in the retention of postparietals, jugals, and (maybe) tabulars.


Albanerpetontidae:
(Jurassic - Neogene) A minor group of extinct lissamphibians. Distinguished by features of cranial osteology, including non-pedicellate three-cusped teeth. Resembling scaly salamanders. For us, their important role is to remind us that the loss of scales in Amphibia only occurred inside Lissamphibia. Thus, we should not assume, as many artists do, that ancient amphibians had naked skin like that of frogs and salamanders.

Up close, the skull of an albanerpetontid looks like a salamander attempting to become a caecilian. The neurocranium is strengthened by the fusion of prootics, opisthotics, and exoccipitals into an otic bone, however the caudate cheek-gap and hinged squamosal are still evident. Ruta and Coates, 2007, find albanerpetontods to be stem-gymnophionans. One clear synapomorphy of albanerpetontids:
- Frontals are fused.
Daza et al., 2020 illuminated albanerpetontid ecology with their description of Yaksha perettii, a mid-Cretaceous member preserved in amber. Yaksha sports an elongate entoglossal process of the hyoid arch analogous to those of chameleons, suggesting that like them, it was a "ballistic" ambush predator that captured its prey by projecting its tongue. Was it just Yaksha, or is this a general feature of Albanerpetontids?

Lissamphibian relationships:

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
What we definitely know:
Batrachia: (Early Triassic - Quaternary) The last common ancestor of anurans and caudates. Synapomorphies include:
- The operculum, a cartilaginous element that couples the stapes to the fenestra ovalis.
- The auditory sensory cells of the inner ear are divided into a papilla basilaris and a papilla amphibiorum, in contrast to other vertebrates who have only a single papilla.
- The complete loss of fishy scales and increased reliance upon cutaneous breathing.
Lissamphibian phylogenetic hypotheses:
During the cladistics era, the monophyly and position of Lissamphibia has been extensively debated. We present here the current majority consensus, but the matter is not fully settled.Potential synapomorphies of Lissamphibia, if monophyletic:
- Significant reduction of skull roof bones. Includes the loss of elements:
- Supratemporals
- Ectopterygoids
- Palatal teeth occur in a row that parallels marginal teeth, especially on the vomer.
- Greatly reduced ribs
- Holospondylous vertebrae.
- Several soft-tissue features
- Drastic metamorphosis where morphological and ecologic transition (e.g. aquatic -> terrestrial) coincide and are compressed into a short time. Witzmann, 2015
{kind=link}
{kind=link}

Pros:
- The association is based on unique synapomorphies such as:
- the presence of two-cusped pedicellate teeth (in which the root and crown are separated by a layer of uncalcified tissue)
- The expansion of the interpterygoid vacuity and loss of the ectopterygoid
- The hypothesis' ready accommodation of new fossil taxa, such as the Early Permian "frogamander" Gerobatrachus (Anderson et al., 2007.), the current candidate for sister taxon to Lissamphibia.
- Its robustness when new fossil character information comes to light. (E.G. the description of the braincase of Eocaecilia by Maddin et al. 2012.)
- The fact that reinterpretations of known fossils and anatomical structures seem to reinforce it. E.G: Maddin and Anderson's 2012 identification of homologies of the caecilian otic capsule with that of frogs and salamanders.
{kind=link}
- A significant morphological gap separates Gymnophiona and everything else in this scheme.

{kind=link}
- It pushes the caecilian record farther back in time.
- The new taxon, while sharing potential synapomorphies with caecilians, appears to be nested in Stereospondyli as a trematosaurian, close to metoposaurs!
Whatever its relationships, the aestivation burrows of Chinlestegophis and its close relative Ninumbeehan dookoodukah are an interesting adaptative response to the megamonsoonal environment of the Carnian Pluvial Episode of the Late Triassic. (So et al., 2024.)

Funcusvermis gilmorei (Andrey Atuchin/National
Park Service/Petrified Forest Museum Association)
So we wait and hope. Monophyletic Lissamphibia within Dissorophoidea remains the front runner for now.
- Jason Anderson, Robert Carroll, Timothy Rowe 2003. New information on Lethiscus stocki (Tetrapoda: Lepospondyli: Aistopoda) from high-resolution computed tomography and a phylogenetic analysis of Aistopoda. Canadian Journal of Earth Sciences, 40(8): 1071-1083
- Jason Anderson 2007. Incorporating Ontogeny into the matrix: A Phylogenetic Evaluation of Developmental Evidence for the Origin of Modern Amphibians. In: J. Anderson and H-D Sues Eds. Major Transitions in Vertebrate Evolution. Indiana University Press, Bloomington and Indianapolis. pp. 182-227.
- Jason Anderson, Robert Reisz, Diane Scott,Nadia Froebischer, and Stuart Sumida, 2007. A stem batrachian from the Early Permian of Texas and the origin of frogs and salamanders. Nature 453, 515-518.
- Robert Carroll, 2009. The Rise of Amphibians: 365 Million Years of Evolution. Johns Hopkins University Press, Baltimore. 360 pp.
- Aja Mia Carter, S. Tonia Hsieh, Peter Dodson, Lauren Sallan, 2021. Early amphibians evolved distinct vertebrae for habitat invasions. PLOS One 16(6): e0251983
- Mariana Chuliver, Federico L. Agnol’n, Agust’n Scanferla, Mauro Aranciaga Rolando, Mart’n D. Ezcurra, Fernando E. Novas, and Xing Xu 2024. The oldest tadpole reveals evolutionary stability of the anuran life cycle. Nature 636, 138Đ142
- Jennifer Clack, 2014. Gaining Ground: The Origin and Evolution of Tetrapods. Indiana University Press, Bloomington. 523 pp.
- Michael Coates, Marcello Ruta, and Matt Friedman. 2008. Ever Since Owen: Changing Perspectives on the Early Evolution of Tetrapods. Annual Review of Ecology, Evolution, and Systematics. 2008. 39:571Đ92.
- Juan Daza, Edward Stanley, Arnau Bolet, Aaron Bauer, Salvador Arias, Andrei Cernansky, Joseph Bevitt, Phillip Wagner, and Susan Evans. 2020. Enigmatic mid-Cretaceous amphibians were chameleon-Like ballistic feeders. Science 370:687-691.
- Nadia B. Fröbisch and Neil H. Shubin. 2011. Salamander limb development: Integrating genes, morphology, and fossils. Developmental Dynamics, 240(5):1087-1099.
- Marc E. H. Jones , Roger B. J. Benson, Pavel Skutschas, Lucy Hill, Elsa Panciroli, Armin D. Schmitt, Stig A. Walsh, and Susan E. Evans. 2022. Middle Jurassic fossils document an early stage in salamander evolution. Proceedings of the National Academy of Sciences, 119 (30) e2114100119.
- Jozef Klembara, Jennifer Clack, Andrew Milner, and Marcello Ruta. 2014. Cranial anatomy, ontogeny, and relationships of the Late Carboniferous tetrapod Gephyrostegus bohemicus Jaekel, 1902. Journal of Vertebrate Paleontology, 34(4):774Đ792.
- Ben T. Kligman, Bryan M. Gee, Adam D. Marsh, Sterling J. Nesbitt, Matthew E. Smith, William G. Parker and Michelle R. Stocker. 2023. Triassic stem caecilian supports dissorophoid origin of living amphibians. Nature 2023.
- Michel Laurin and Robert Reisz. 1997. A new perspective on tetrapod phylogeny. In: S. Sumida and K. Martin eds. Amniote Origins. Academic Press. New York. 9-59.
- Hillary Maddin and Jason Anderson. 2012. Evolution of the Amphibian Ear with Implications for Lissamphibian Phylogeny: Insight Gained from the Caecilian Inner Ear. Fieldiana Life and Earth Sciences. 5 :59-76.
- Hillary Maddin, Farrish Jenkins, Jason Anderson. 2012. The Braincase of Eocaecilia micropodia PLOS One, December 2012. DOI: 10.1371/journal.pone.0050743.
- Pedro Luiz Mailho-Fontana, Marta Maria Antoniazzi, Cesar Alexandre, Daniel Carvalho Pimenta, Juliana Mozer Sciani, Edmund D. Brodie, Jr., and Carlos Jared. 2020. Morphological Evidence for an Oral Venom System in Caecilian Amphibians. CellPress 23(7) July 4, 2020.
- Arjan Mann, Jason D Pardo, and Hans-Dieter Sues. 2022. Osteology and phylogenetic position of the diminutive ÔmicrosaurŐ Odonterpeton triangulare from the Pennsylvanian of Linton, Ohio, and major features of recumbirostran phylogeny. Zoological Journal of the Linnean Society 2022;, zlac043,
- David Marjanovic and Michel Laurin. 2019. Phylogeny of Paleozoic limbed vertebrates reassessed through revision and expansion of the largest published relevant data matrix. PeerJ 6:e5565
- David Marjanovic, Hillary C Madden, Jennifer Lori, and Michel Laurin. 2024. The new problem of Chinlestegophis and the origin of caecilians (Amphibia, Gymnophionomorpha) is highly sensitive to old problems of sampling and character construction. Fossil Record 27(1) 55-99
- Claudia A. Marsicano, Jason D. Pardo, Roger M. H. Smith, Adriana C. Mancuso, Leandro C. Gaetano, and Helke Mocke. 2024. Giant stem tetrapod was apex predator in Gondwanan late Palaeozoic ice age. Nature 631, 577-582
- Angela Milner. 1988. The relationships and origins of living amphibians. In: M. Benton ed. The phylogeny and classification of tetrapods, volume I: amphibians, reptiles, birds. Clarendon Press. Oxford. 59-102.
- Darren Naish. 2013. Tetrapod Zoology blog - Teenage Mutant Ninja Temnospondyls. retrieved from http://blogs.scientificamerican.com/tetrapod-zoology/2013/09/24/teenage-mutant-ninja-temnospondyls/, February 2015.
- Pardo, J. D., Small, B. J., Huttenlocker, A. K. 2016. A caecilian-like temnospondyl from the Triassic Chinle Formation of Colorado and its bearing on the origins of Lissamphibia. Presentation at 76th Annual Society of Vertebrate Paleontology meetings.
- Jason D. Pardo, Matt Szostakiwskyj, Per E. Ahlberg & Jason S. Anderson, 2017. Hidden morphological diversity among early tetrapods. Nature 546, 642Đ645.
- Pardo, J. D., Small, B. J., Huttenlocker, A. K. 2017. Stem caecilian from the Triassic of Colorado sheds light on the origins of Lissamphibia. Proceedings of the National Academy of Sciences 114 (27).
- Laura B. Porro,Emily J. Rayfield, and Jennifer A. Clack. 2023. Computed tomography and three-dimensional reconstruction of the skull of the stem tetrapod Crassigyrinus scoticus Watson, 1929. Journal of Vertebrate Paleontology, 42(4). https://doi.org/10.1080/02724634.2023.2183134.
- Yu-Fen Rong, Davit Vasilyan, Li-Ping Dong, and Yuan Wang. 2020. Revision of Chunerpeton tianyiense (Lissamphibia, Caudata): is it a cryptobranchid salamander?. Paleoworld Pre-proof
- Marcello Ruta and Michael Coates. 2007. Dates, nodes and character conflict: Addressing the Lissamphibian origin problem. Journal of Systematic Paleontology, 5:1, 69-122
- Rainer Schoch. 2013. The evolution of major temnospondyl clades: an inclusive phylogenetic analysis. Journal of Systematic Paleontology, iFirst 2013, 1-33
- Rainer Schoch. 2018. The putative lissamphibian stem-group: phylogeny and evolution of the dissorophoid temnospondyls. Journal of Paleontology, 93(1)
- Rainer Schoch and Florian Witzmann. 2024. The evolution of larvae in temnospondyls and the stepwise origin of amphibian metamorphosis. Biological Reviews 99(5):1613-1637
- Calvin So, Aaron M. Kufner, Jason D. Pardo, Caian L. Edwards, Brandon R. Price, Joseph J. Bevitt, Amanda LeClair-Diaz, Lynette St. Clair, Josh Mann, Reba Teran and David M. Lovelace. 2024. Fossil amphibian offers insights into the interplay between monsoons and amphibian evolution in palaeoequatorial Late Triassic systems. Zoological Journal of the Linnean Society B. 291(2033)
- Matt Szostakiwskyj, Jason D. Pardo, Jason S. Anderson. 2015. Micro-CT Study of Rhynchonkos stovalli (Lepospondyli, Recumbirostra), with Description of Two New Genera. Plos|One June 10, 2015.
- Florian Witzmann. 2015. CO2-metabolism in early tetrapods revisited: inferences from osteological correlates of gills, skin and lung ventilation in the fossil record. Lethaia 49(4), 492Đ506.
- Jason Anderson 2007. Incorporating Ontogeny into the matrix: A Phylogenetic Evaluation of Developmental Evidence for the Origin of Modern Amphibians. In: J. Anderson and H-D Sues Eds. Major Transitions in Vertebrate Evolution. Indiana University Press, Bloomington and Indianapolis. pp. 182-227.