GEOL 331 Invertebrate Paleontology
Fall Semester 2008
Vertebrate Paleontology
|
Aquatic Vertebrates
"Ostracoderms": The proper fossil record of vertebrates begins with a huge paraphyletic grade of armored jawless vertebrates. All share
with gnathostomes:
- Mineralized exoskeleton (i.e., a dermal skeleton), including the hard tissues dentine,
bone, dentine, and enamel
- Sensory-line canals and grooves
|

|
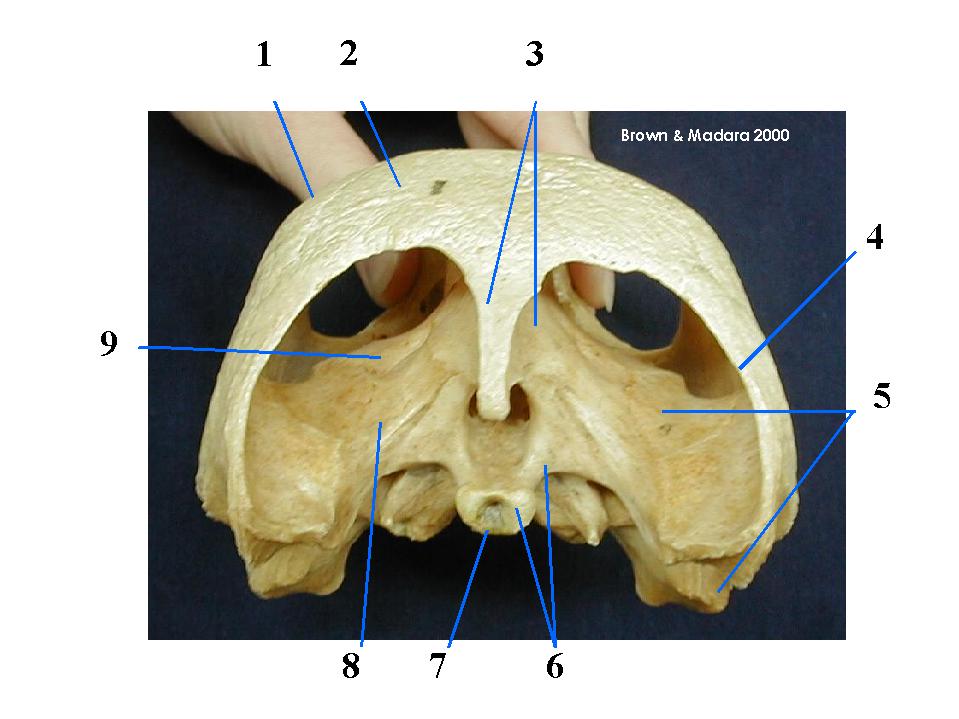
A rogue's gallery of fossil jawless vertebrates
|
With euconodonts and ganthostomes possessing hard tissues made of calcium phosphate. Conodonts were not the only representation of craniate hard tissues in the Cambrian, however. Enigmatic, scale-like plates of bony armor called Anatolepis were also present.
|
|
Fairly complete fossil skeletons of creatures with Anatolepis-like bone first appear in the Ordovician in the form of Sacabambaspis.
In this creature, all bone takes the form of flat plates armoring the external surface. It seems to have played the roles of:
- Armor
- A reservoir of calcium and phosphate for use by the animal's metabolism.
It was not an internal skeleton. This bone resembles the Dermal bone - formed by the ossification of a membrane in living animals. Dermal bones tend to be flat and plate-like. E.g. the bones of the top of the human braincase.
|
Pteraspidomorphi (Cambrian (assuming Anatolepis) - Devonian). By the Silurian, many Sacabambaspis - like creature are known.
Morphology:
- in the form of acellular dermal armor. We see two armor shields, one dorsal and one ventral. Each gill opening is protected by an individual bone plate. All of this bone is dermal and forms external armor over the creature. It has no internal bone.
- The mouth is opened and closed by row of narrow parallel plates in the lower "lip."
- A head with small eyes and otic capsules above the mouth.
- Impressions of otic capsules on the underside of the dorsal shield indicate the presence of two sets of semicircular canals - similar to lampreys.
- Caudal fin is either diphycercal or hypocercal.
|

|
Without a living analog, speculations about the ecology of these creatures is very limited. We note the following:
Feeding:
- If the distribution of pharyngeal slits is any indicator, they had a substantial pharynx.
- They lacked jaws, or even the keratinous structures that hagfish and lampreys use to process food. The implication was that they were somehow filtering particulate matter. General speculation is that they were suspension feeders: They lived like ammocoetes, resting on or in the bottom and suspension feeding from the water - seems more likely given that their oral plates never seem to suffer from abrasion or wear..
Locomotion:
- Pteraspidomorphs lacked the surfaces that modern aquatic vertebrates use to control their direction in the water. They could, at best, propel themselves Branchiostoma-style.
- Experiments with models indicate that when not being actively propelled, the bodies of pteraspidomorphs would settle onto the substrate without diving or rolling.
Leads to the general picture of pteraspidomorphs as creatures that rested on the bottom to suspension feed, but would swim haphazardly when disturbed, only to settle to the bottom again. Their probably poor control over swimming orientation and direction.
|
Anaspida (Silurian)
Morphology:
- in the form of acellular scales. No significant armor
- Body is cylindrical with notochord supporting lower lobe of caudal fin.
- Pharyngeal (gill) openings are small and form a short, posteriorly slanting row.
- While there are no paired fins, there are paired triangular spines in a vaguely "pectoral" position.
|

|
|
Overall, anaspids seem adapted for active swimming. exactly how they ate is mysterious, but they lack the obvious adptations to suspension feeding of pteraspidomorphs. Phillipe Janvier has argued persuasively that anaspids might be the sister taxon to lampreys (Hyperoartia). That would make the lampreys' lack of hard tissue an evolutionary reversal.
|
>
|
|
Thelodonti (Ordovician - Devonian)
Morphology:
- Entirely consists of small scales that usually disarticulate when the animal dies. These scales are distinctive, consisting of enamel and dentine layers around a pulp cavity, like a vertebrate tooth. Note: These are new types of bone tissue:
- Dentine: Dense acellular bone
- Enamel: Hard tissue consisting entirely of interlocking crystals of calcium phosphate - the mineral component of bone.
- In some cases, these tooth-like scales line the oral cavity and mouth.
- Again, no paired fins, there are paired triangular "fin flaps" without skeleton or muscle.
- Hypocercal caudal fin.
|

|
Endochondral bone: In all of the above taxa, there is no consensus about their interrelationships or closeness to jawed vertebrates. But then we get two groups that share a major synapomorphy with gnathostomes: Endochondral bone is preformed in cartilage, instead of being laid down directly onto a membrane. This fact allows endochondral bone to assume complex three dimensional shapes. Endochondral bone makes its evolutionary debut in the skeleton of the braincase.
|
Galeaspida: Restricted to southern China and Indochina, then a separate continent. (Silurian - Devonian)
Morphology:
- large flat head shield enclosing an endochondral bony braincase.
- large opening in upper front of head shield leads to olfactory capsules and pharynx
- mouth is ventral
- Pharynx is large with many gill openings, suggesting suspension feeding.
- No suggestion of paired fins.
- Overall, although the equipment is different, the life-style looks like that of pteraspidomorphs.
|

|
|
Osteostraci (Silurian - Devonian): Resemble galeaspids but with differences:
Morphology:
- Head shield contains large sensory fields.
- Head shield fused with endochondrally ossified braincase.
- Notochord invades upper lobe of tail
- Dorsal fin
- But the big thing: Paired pectoral fins (complete with endochondral skeleton elements and muscle) present.
|

|
Phylogenetically, this diversity is problematic. A few potential synapomorphies include:
- Paired nostrils: Ptersapidomorphi, Thelodonti, Gnathostomata
- Presence of mineralized tissue: All groups, including Euconodonta, but excluding Hyperotreti (hagfish) and Hyperoartia (lampreys)
- Annular cartilage: Hyperoartia and Anaspida
There is no consensus on the phylogeny of these creatures, except for two good synapomorphies:
- Endochondral bone: Galeaspida, Osteostraci, Gnathostomata
- Paired pectoral fins: Osteostraci, Gnathostomata
All are interesting and specialized creatures, but as long as jawless, gills had to serve "double duty": as organs of suspension/deposit feeding and of
respiration.
Gnathostomata: (Sil - Rec.) The jawed vertebrates. Quantum leap forward.
|
Synapomorphies
|

|
Origin of jaws: Many gnathostome synapomorphies are continuations of longstanding vertebrate trends toward increased skeletal ossification, brain enlargement, and improvement of swimming. Big exceptions are changes to the pharynx, especially jaws. We have:
- Jaws themselves - palatoquadrate and meckel's cartilage
- Internal branchial skeleton.
- Inductive formation of pharyngeal calcifications.
Three general hypotheses account for this radical transformation:
Serial homology and Carl Gegenbaur: Gegenbaur in 1870 proposed that the jaws and hyoid arch represented modified anterior branchial arches. In many modern gnathostomes, the jaws and hyoid arch enclose an opening called the spiracle (=eustacean tube in land verts) Originally, in Gegenbaur's scenario, they originally would have been separated by a complete gill slit. Gegenbaur predicted that fossil forms would be found with this configuration, but after a century, this has not happened.
New Mouth: Jon Mallatt, in the 1990s, proposed that the jaws and hyoid arch developed from arches further back in the branchial array. Thus, the "old mouth" (like the oral cavity of Branchiostoma) is homologous to the space between the teeth and lips, while the oral cavity posterior to the "new mouth." This tied up one loose end, the presence of labial cartilages in sharks. Still subject to paleontological objections that beset Gegenbaur's scheme.
Neomorphic jaws: Mid 1970s, Bobb Schaeffer of AMNH proposed that jaws were neomorphs, inductively derived from the interaction of endo and ectodermal tissue when modifications of the forebrain brought the pharyngeal endoderm and neural tube ectoderm into contact. This has the advantage of explaining the absence of intermediate forms and the simultaneity of the appearance fo jaws and the increase in brain size Possible cause - HOX gene duplication increasing the size of the brain.
The Velar skeleton: In 1996, Phillippe Janvier proposed that the skeleton of the velum (as seen in lampreys) was the precursor to the mandibular arch. Like jaws, the cartilagenous velar elements of lampreys have upper and lower segments articulating at a posteriorly facing hinge. Alas, here, too, paleontological evidence currently falls short, as it it not obvious that other jawless vertebrates had a velar skeleton. Of course, such a structure would be hard to preserve.
Gnathostome diversity: Three major groups:
- Placodermi
- Chondrichthyes
- Osteichthyes.
Placodermi: Silurian to Devonian armored gnathostomes experienced a rapid worldwide diversification and sudden decline.
Placoderms were very diverse and occupied a wide range of ecological roles. Their specializations included:
Fossil record: Fragmentary records of placoderms appear in the Middle Silurian. this is followed by a rapid diversification. During the Devonian, placoderms were the dominant vertebrate group. Both marine and fresh water forms are recorded with a worldwide distribution except for puzzling absence in South American sediments. Placoderm diversity was greatly reduced by an extinction event in the Late Devonian. They were completely extinguished by the mass extinction event at the end of the Devonian. Thus, entire radiation took up only about 50 million years, but while it lasted, it was spectacular.
|
What placoderms lacked:
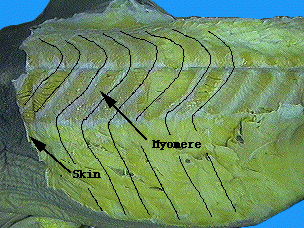
The myomeres of living gnathostomes differ from those of lampreys and hagfish in that there myomeres are divided into upper and lower halves by a horizontal septum of connective tissue. Lampreys' are not. Recent work on placoderm fossils with soft tissue preservation show that the placoderms, like lampreys, lacked the horizontal septum. This feature, therefore, is a synapomorphy of the two remaining gnathostome groups: Chondrichthyes and Osteichthyes.
|

|
|
Chondrichthyes: (Sil - Rec.)
Sharks, chimaeras, and relatives.
- Synapomorphies:
- Skeleton calcifies via small apatite prisms rather than ossifying as bone.
- Dermal skeleton consisting of small denticals and plates.
- Denticle morphology: neck and basal canals . Sound familiar? Denticles are histologically like teeth.
- Unique tooth replacement pattern. (Relatively slow in primitive chondrichthyan, accelerating in later ones.)
|

Cladoselache: Late Devonian.
|
Sampling of diversity:
- Cladoselache: Late Devonian. Relatively primitive and superficially shark-like.
- Holocephali: Chimaeras and fossil relatives
- Elasmobranchii: Shark-like chondrichthyans.
People speak of sharks as being primitive, but living elasmobranchs are all highly derived with respect to their Paleozoic (Orthacanthus) and Mesozoic (Hybodus) relatives. NOTE: Rays and skates are elasmobranchs that have become secondarily adapted for bottom dwelling.
Fossil record: First well documented species from Late Devonian, however shark-like denticles known from Silurian. Explosion in diversity in early Penn, followed by decline. Increase in diversity in Jurassic, and explosion in Neogene. Max diversity in Miocene.
|
Osteichthyes: bony fish. (Sil - Rec)
- Synapomorphies:
- Unique pattern of dermal bones of head and pectoral girdle. (clavicle, interclavicle, cleithrum, parasphenoid, etc.)
- Dermal skull bones with internal descending laminae that interact with endochondral elements.
- Branchiostegal rays present: These are the narrow elements that cover the throat and part or all of the gill covers.
- Lungs: The presence of lungs - outpouchings of the esophagus used to obtain suplimentary oxygen from "swallowed" air. As we will see, Actinopterygii and Sarcopterygii exploit this synapomorpy in very different ways.
- Operculum: A large plate of dermal bone suspended from the hyoid arch covering and protecting the gill arches. (Ancestrally consists of three bones, the preopercular, opercular and subopercular.)
|

Strunius: Late Devonian.
|
Major groups:
- Actinopterygii: Ray-finned fish
- Sarcopterygii: Lobe-finned fish
Psarolepis (Early Devonian) is our closest well-known approximation of the last common ancestor of Osteichthyes.
This creature has features seen in both primitive sarcopterygians and actinopterygians, but retains others typical of more primitive gnathostomes, such as the stout pectoral spine.
|
Actinopterygii: (Dev - Rec)
The vast majority of "fishlike" bony fish.
- Identification:
- Diversity: Immense
- A Sampling of primitive and derived groups.
- Cheirolepis: (Dev.) Most primitive well-known actinopterygian.
- Cladistia: Reedfish or bichirs. Fossil record from Cretaceous but lineage must extend from Devonian. Most primitive living.
- Chondrosteans: Sturgeons, paddlefish, and extinct saurichthyans. (Tri - Rec.)
- Ginglymodi: Gars. (K - Rec.)
- Halecomorphi: Amia (Bowfin) and relatives. (Tri - Rec.)
- Teleostei 99% of actinops. (Tri - Rec.)
|

Cheirolepis: Late Devonian.
|
|
Sarcopterygii:
The lobe-finned fish and tetrapods.
|

Osteolepis: Middle Devonian.
|
Primitive sarcop diversity:
- Actinistia: Coelacanths. (Dev - Rec)
- Synaps:
- Diversity: Very diverse in Late Paleozoic, hit hard by P extinction. In Mesozoic, two lineages apparant, one of large predatory fresh water specialists, one of marine fish leading to Latimeria. Disappear in Cretaceous.
- Discovery of Latimeria revealed function of bipartite cranium as an adaptation for delivering a stronger bite.
|

Latimeria
|
- Choanata: (Dev - Rec) Choanae.
- Dipnoi: (Dev - Rec) Lungfish and relatives.
|

African lungfish Protopterus anectens
|
|
Tristichopteriformes: A group of choanate fish ranging from the Devonian to the early Permian. they are interesting to us as indicators of what the last fully aquatic ancestors of land vertebrates might have looked like. Fortunately, the anatomy of some, such as Eusthenopteron, is very well know from many excellent fossils.
- Fossils found in fresh water deposits. Their disproportionately small eyes compare well with living fish that inhabit muddy bodies of water. In that realm, they were probably powerful predators.
- It is often suggested that they were able to move on land. Although it is conceivable that they might have dragged themselves to new bodies of water (evidence includes the roughly equal length of radius and ulna in some) it seems clear from their general profile that this would be an emergency procedure.
|

Eusthenopteron fordii from
Olduvai George
|
|
Panderichthyes: This is an example of a small group of fish-like vertebrates even closer to the common ancestry of land vertebrates than Osteolepiformes:
- Synapomorphies with land vertebrates:
- Complete loss of dorsal fins.
- Frontal bone in dermal skull roof
- Eye sockets located on the dorsal surface of the skull.
- Biological interpretation: Panderichthyes, although still aquatic, was anatomically somewhat specialized for shallow water or vanishingly brief excursions onto land.
|

|

|
Tiktaalik (Late Devonian):
First published in 2006, this creature has become the new poster-child of vertebrate evolution. It is featured in:
In its general profile, it's similar to Panderichthyes, there are two important differences.
- The wrist. Although fin rays are still present (and digits absent), its wrist is capable of flexing. Thus, it could support at least some of its weight on its forelimbs. No indication that it ever emerged completely from the water.
- Its skull and pectoral girdle were no longer in contact, allowing the head to be flexed with respect to the torso - the beginnings of the neck.
Tiktaalik's pelvis and hind limbs have not been reported formally, but preliminary reports suggest that they are surprisingly robust.
|
Acanthostega (Latest Devonian):
Very well known from relatively complete skeletons.
- Breathing
- Open operculum and functional gills present in adult.
- Notch in rear of skull apparantly accomodated fish-style spiracle, not an impedance-matching ear.
- Mechanical support
- Flexible wrist, ankle
- Slightly flexible elbow and knee.
- Fin rays lost from all but the caudal fin
- Fingers (eight per limb) and toes (seven per limb) present.
- accessory articulations between neural arches, but no zygapophyses to speak of.
- Pelvis is small but has a ligamentous connection with a sacral rib.
- Feeding:
- Skull and pectoral girdle detached.
- First vertebra is enlarged, but otherwise no specialization of the vertebral column, such as zygapophyses.
- Senses:
- Extensive lateral line system
- Robust hyomandibula/stapes articulates with fenestra ovalis.
- Overall impression is of an aquatic animal that perhaps primarily used hands and feet in walking on bottom or through underwater vegetation.
To Next Lecture.
To Previous Lecture.
To Syllabus.









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}