

Vertebrata II: Tetrapoda
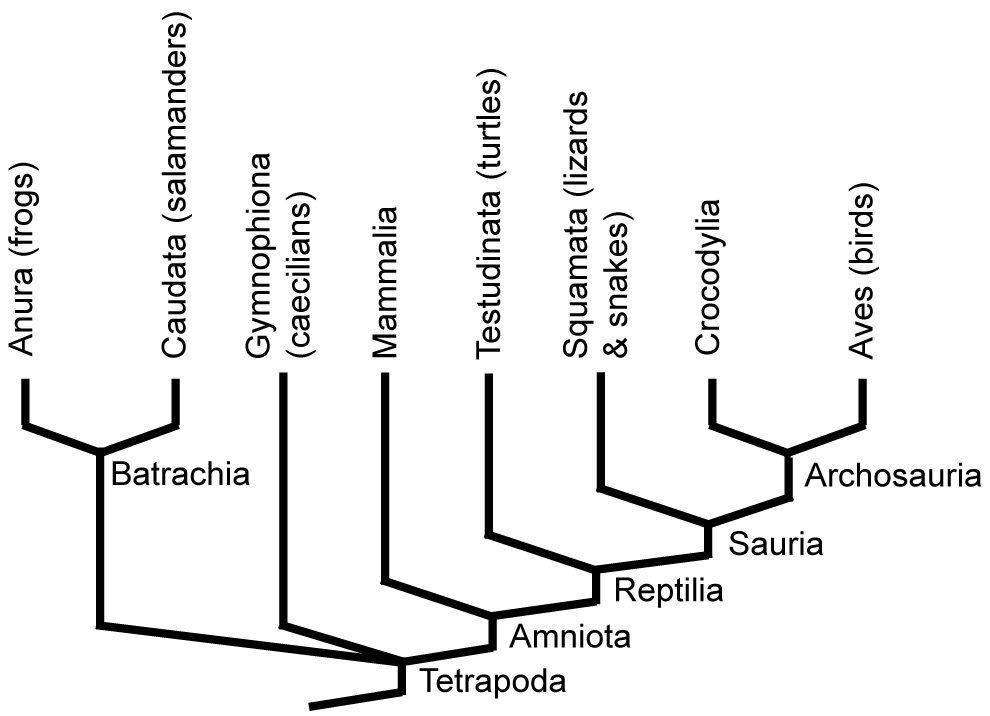
Extant tetropod phylogeny
Last lecture, we saw the diversity of marine vertebrates ("fish"), ending with the transition from fully aquatic forms to taxa possessing features that could be exapted for life on land. Now we look at the invasion of the land and the diversity of Tetrapoda, the descendants of the vertebrates that accomplished this. Major living tetrapod groups include:
- Anura: (Triassic - Holocene) Frogs.
- Caudata: (Jurassic - Holocene) Salamanders.
- Gymnophiona: (Jurassic - Holocene) Caecilians.
- Mammalia: (Jurassic - Holocene) Mammals.
- Testudinata: (Permian - Holocene) Turtles.
- Squamata: (Triassic - Holocene) Lizards (including snakes).
- Crocodylia: (Triassic - Holocene) Crocodilians.
- Aves: (Cretaceous - Holocene) Birds.
The tetrapod evolutionary story is interesting for reasons going far beyond their patten of diversity. Above all, at several stages of their evolution, circumstances combined to push then through profound biomechanical transformations associated with life on land. Foremost are:
- Conquest of the shoreline: the ability to emerge from the water to feed and thermoregulate.
- Conquest of land: The ability to spend substantial time out of water as adults
- The rise of the cleidodic egg: The ability to reproduce on land
Roster of transformations.
- Breathing: The ability to breathe air to some degree was inherited from the common ancestor of Osteichthyes. The problem with life on land wasn't the absence of lungs. Rather it was the presence of gills, which became a true liability on land. If you spent any great time out of water, you would dehydrate by losing water through the gills. Thus, gills were retained only by creatures that still spent the bulk of their time in the water at all life stages and only ever emerged briefly. We can't usually see gills directly in fossil organisms, but we can infer their presence from features of the skeleton near them.
- Mechanical support: In water, one's weight is supported by bouyancy, and the limbs function in propulsion and control of orientation. On land, the limbs took over the function of supporting the body. Thus:
- Paired limbs became larger, more robust, and more flexible. Remember, a tristichopterid, whose radius was twice the length of its ulna, could not have bent its arm effectively.
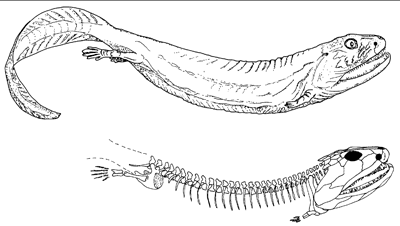
- Digits: In contrast to every other vertebrate this course has deal with, most members of Stegocephali lack fin rays on their paired fins and, instead, have, digits - fingers and toes. These first appear in animals that are otherwise clearly aquatic. The mechanism of their appearance is enigmatic, but they seem at first to have been used as a means of walking on the bottom or pushing through aquatic vegetation.
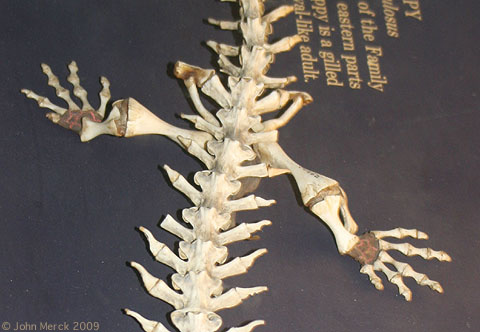
Andrias japonicus (Japanese giant salamander) - Sacrum: A bony attachment was established between the pelvis and vertebral column, enabling the hind limbs to support the weight of the torso.

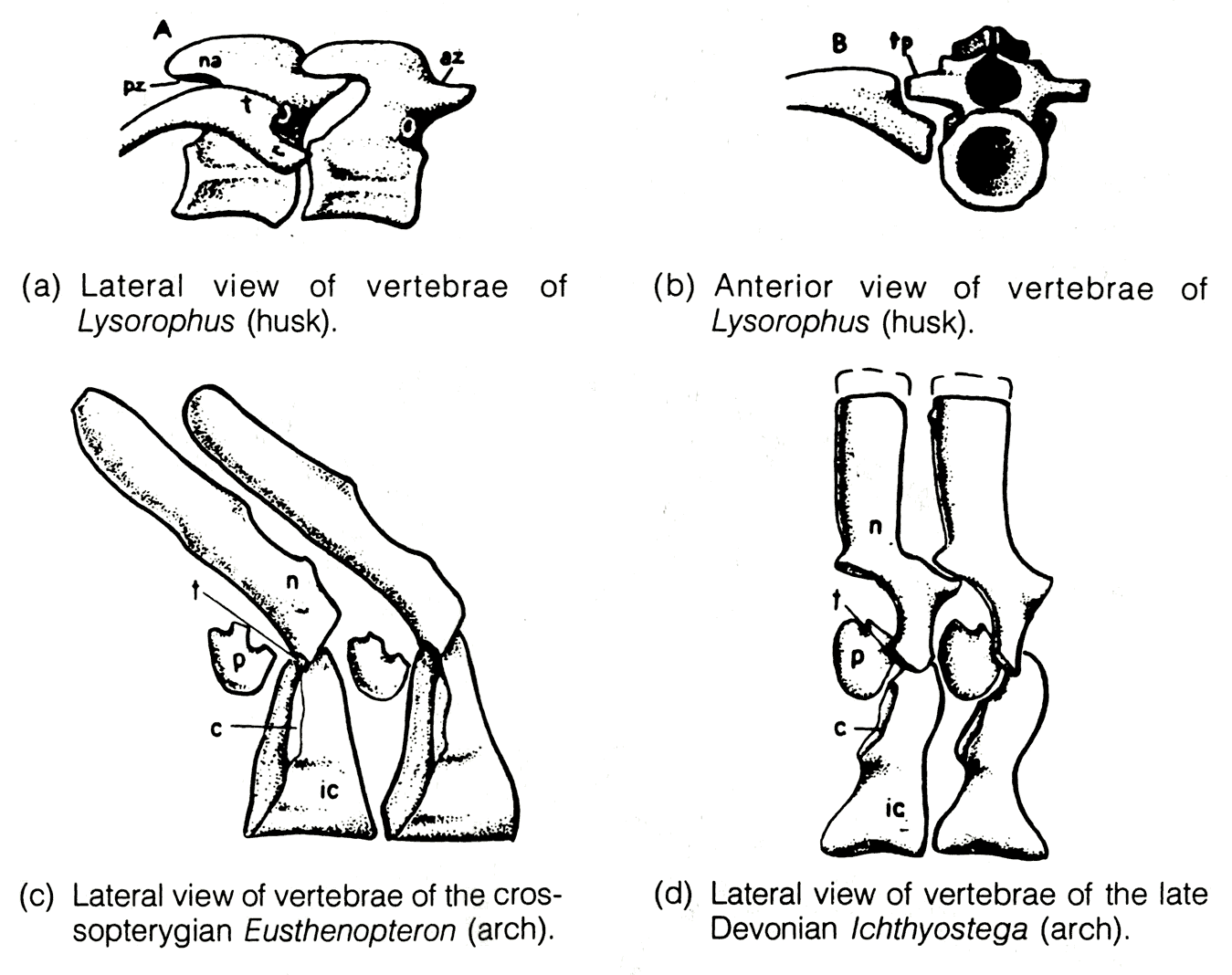
from Systematic Biology- Vertebral centra: These were small or absent in aquatic sarcopterygians, which retained an exposed notochord. Movement on land demanded that the notochord be strengthened against bending and compression. Thus, robust centra formed as ossifications of the notochord.
- Feeding: A fish does not need to bend its neck to pick up food from the ground. It just swims up to it and bites. On land, the head must move relative to the torso. Result is a neck.
- In an ancestral choanate like Eusthenopteron, the pectoral girdle is effectively part of the head, united to it by a series of dermal bones. The evolution of the neck requires the loss of these bones.
- Senses: All of the senses must be retooled to function in air.
- Vision: Air and water have different refractive indices, such that one's lens must be adapted to one or the other. The position to the eye sockets in Panderichthys suggest that the ability to see in air could have evolved long before the ability to walk on land.
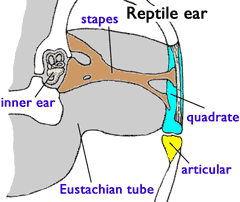
- Hearing: The density of the body is similar to that of water. Thus, sound can pass directly from water through the body into the otic capsule at full intensity. Air is much less dense than water. Thus, for airborn sound to register in the otic cpasule, it must be collected and amplified. This is accomplished by an impedance matching ear:
- Sound is collected over a broad area by a tympanum (A.K.A. ear drum) or through the jaw.
- In land vertebrates, a rod-like bone called the stapes transmits the vibrations of the tympanum to a small opening in the otic capsule, the fenestra ovalis (oval window.) By being concentrated into a small area, the sound is amplified so that it can be detected by the inner ear.
- Where does the stapes come from? It is a modified hyomandibula. The air filled space it occupies is called the middle ear. Together with the eustacean tube that connects the middle ear to the pharynx, it is homologous to the spiracle of aquatic vertebrates.
As we will see, the impedance-matching ear evolved slowly and independently several times in different lineages.
- Sound is collected over a broad area by a tympanum (A.K.A. ear drum) or through the jaw.

Arapaima with conspicuous lateral line- Lateral line sense: Useless out of water. Still, as an ancestral feature, they tend to be retained by adults that spend any significant amount of time in the water.
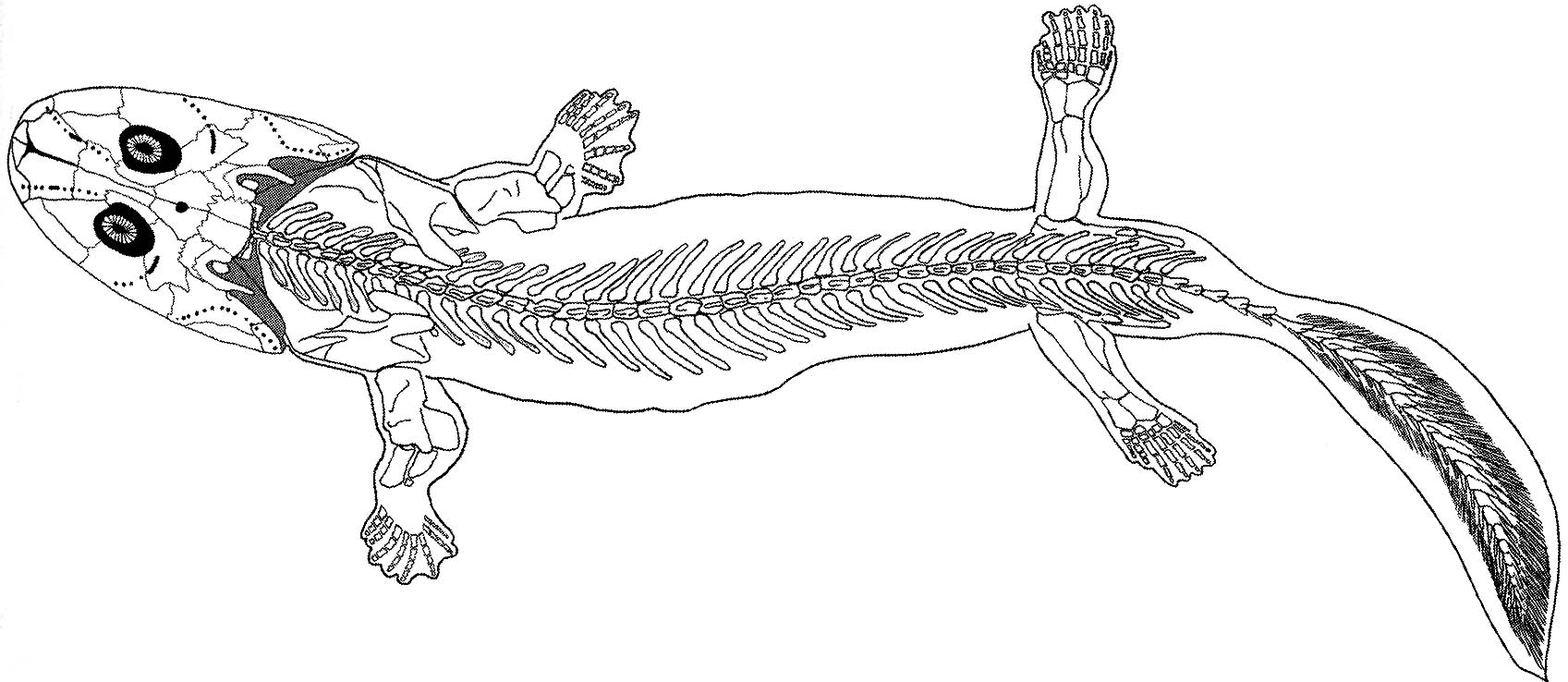
Stegocephalia: The Fingered Vertebrates (Late Devonian - Holocene).Relative to their kin, stegocephalians (tetrapods and their extinct relatives). Acanthostega (right) is representative of their basal condition. Synapomorphies:
- A reduction in the number of skull bones
- A neck (i.e., separation of the skull from the pectoral girdle)
- Fin rays replaced by (recent evidence shows "modified into") segmented bony digits (aka fingers and toes)
- At least one sacral vertebra: that is, the pelves contact at least one vertebra
Romer's Gap - a geologic enigma: Acanthostega lived roughly 365 mya. The earliest known members of Tetrapoda (last common ancestor of living tetrapods) are from roughly 335 mya. This gap - Romer's Gap - coincides with the Hexapod Gap in the insect record. Even after Romer's Gap ended, most stegocephalians remained aquatic with weak limbs and prominent lateral line systems. Examples:
- Greererpeton
- Spathicephalus: With a flat head and teeth specialized for underwater suspension-feeding.
- Crassigyrinus
All of these show ancestral characters indicting aquatic habits as adults:
- Large load-bearing hyomandibula/stapes.
- Extensive lateral line systems.
- Under-sized, weak limbs.
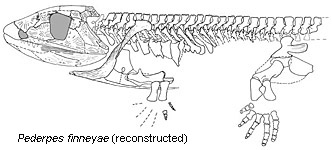
Conquest of the shoreline: In 2002, Jennifer Clack described Pederpes finneyae, from the middle of Romer's Gap at 350 mya. Although it retains a tiny sixth finger, has a robust lateral line system and heavy hyomandibula/stapes; it also has robust limbs and a skull that is compressed from side-to-side like early land vertebrates. We don't know how much time it spent on land, but it seems, at least, to have been able to walk. Recent reports of healed broken limb bones in Pederpes suggest that it spent enough time on land that injury from falls was a danger.
Tetrapoda: For our purposes, the last common ancestor of living amniotes and amphibians and all of its descendants. Synapomorphies include:
- A muscular tongue with glands (although a few living amphibians have a more fish-like tongue)
- The vomeronasal organ, for sensing chemicals left of the ground
- Loss of the internal gills
- Evolution of a new set of external gills in larvae
Tetrapoda breaks down into two total-groups:
- Amphibia: All tetrapods more closely related to frogs and salamanders.
- Reptiliomorpha: All tetrapods more closely related to amniotes.
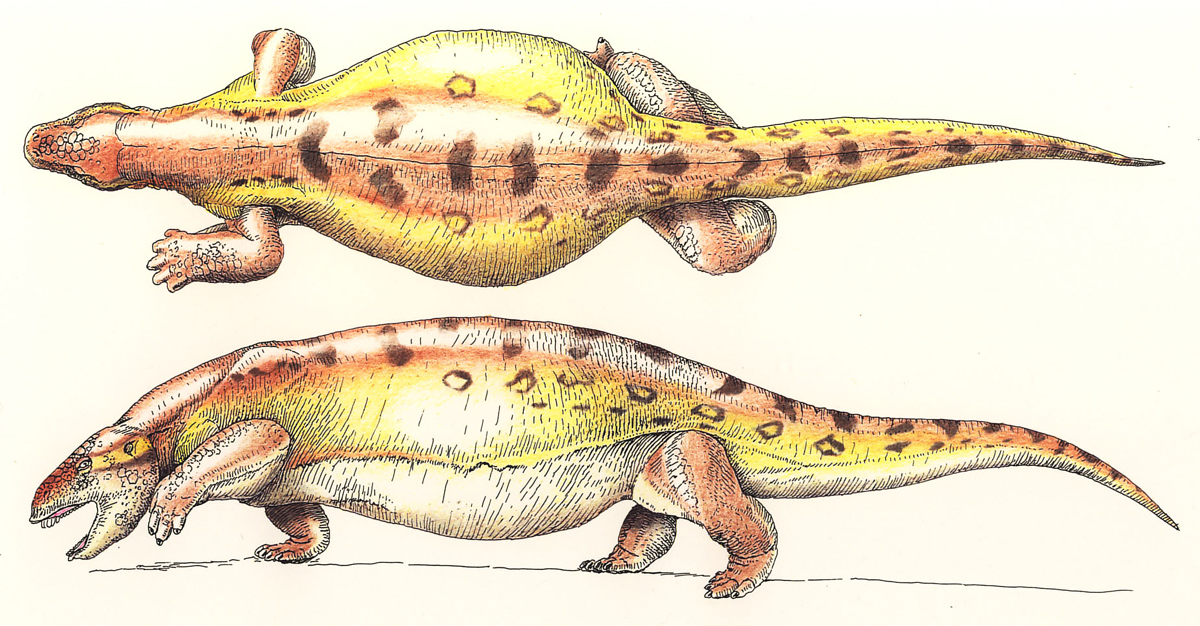
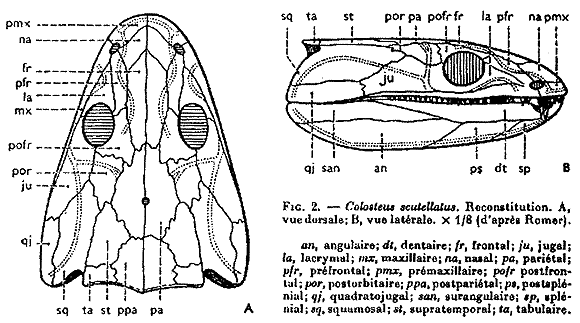
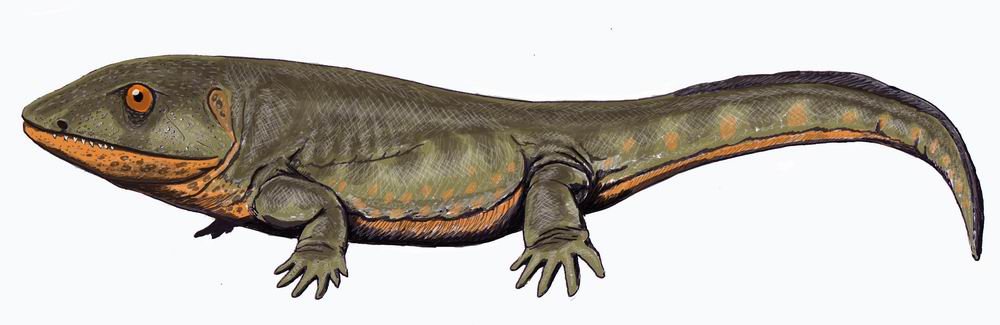
Amphibia: For now, we consider Amphibia. A basal member of this group like Dendrerpeton (right) does not look terribly different form a basal reptiliomorph like Caerorhachis (below) however the "amphibian" lineage acquired certain derived features early on, including:- The opening for the spiracle has become a wide embayment in the rear of the skull roof and the stapes is relatively slender. We interpret that the spiracular opening is closed by the a tympanum and that, together with the stapes, it functions in an impedance-matching ear. (The first of several such derivation.)
- A four-fingered hand.
- Wide openings in the palate.
- Large fangs in the palate.
Most fossil "amphibians" belong to a large group called Temnospondyli that is probably paraphyletic with respect to Lissamphibia - the crown group of amphibians. This is a diverse group of tetrapods ranging from the Early Carboniferous to the Cretaceous, although their heyday was the late Paleozoic and earliest Mesozoic. They lived in or near a variety of fresh water and shallow marine aquatic habitats and exhibited a wide range of body forms, some being stout with broad heads, some flattened, and some elongate. Head shapes, likewise varied from long snouted to broad headed forms. All were aquatic predators of some stripe or another. Dendrerpeton is a basal member of this diverse group. It is less than a meter in length, but many temnospondyls achieved the size of crocodiles, and displayed varying degrees of aquatic or terrestrial specializations.
(Want to know more? For all your temnospondyl needs, check out the temnospondyl entries on Darren Naish's wonderful Tetrapod Zoology blog.)
It is only at this level in stegocephalian evolution - well within the amphibian tree - that we can firmly say that we are dealing with a group containing animals that could routinely make prolonged excursions out of water. Still, depositional settings make it clear that even adults of many temnospondyls were primarily aquatic animals.
There is general, though not unanimous agreement that Anura (frogs) and Caudata (salamanders) for a clade Batrachia within Amphibia. Their sister taxon is Gerobatrachus (Permian.) Members of Batrachia are:
- Scaleless: dermal scales inherited from fishy ancestors persist in most fossil tetrapods. Among Anura and Caudata, their absence enables the skin to function as a respiratory organ.
- Significantly reduced ossification of skull roof. Both number of bony elements and their absolute quantity are reduced.
Reptiliomorpha - Closer to Amniota:
Basal reptiliomorphs like Caerorhachis (Early Carboniferous - right) possess evolutionary novelties that characterize Reptiliomorpha, despite outwardly resembling basal amphibians:- The elements of the skull roof and cheek are loosely connected in a mobile articulation.
- In the vertebral column, the ossification of the centra follows a different pattern.
- The tabular bones (forming the corners of the skull roof) contact the parietals. (E.G.: Silvanerpeton. For an example of the amphibian condition see Balanerpeton.)
Like basal amphibians, basal reptiliomorphs show varying degrees of adaptation for life on land, and a range of adaptations, but none are absolutely independent of water. Some interesting highlights that are successively closer to crown-group Amniota:
- Seymouriamorpha: (Permian) Terrestrial adults with:
- Stocky bodies and short tails, indicative of adult life on land
- Impedance-matching ears in embayments at the rear of the skull.
- Very large adults incorporate a second sacral rib and lose grooves in skull for lateral line system.
- Lepospondyli: (Carboniferous - Permian) Small tetrapods with solidly coosified (fused) centra and neural arches and reduced skull roof bones in comparison to ancestral forms. (Where else have we seen this?). Major lepospondyl groups:
- Aistopoda: (Carboniferous) Superficially snake like, elongate, and limbless. Significant reduction of bones of skull roof.
- Nectridia: (Carboniferous) Long, with short limbs and aquatic, often with bizarre cranial modifications, such as the boomerang - headed Diplocaulus. Note: resting trace fossils left by these animals indicate that the "horns" were connected to the body by a fold of flesh. Sorry, artists.
- Microsaurs: (Carboniferous - Permian) An ecologically diverse variety of small critters, ranging from pedomorphic aquatic forms that retain gills throughout life to forms that, based on limb and body proportions, seem primarily terrestrial.
- Aistopoda: (Carboniferous) Superficially snake like, elongate, and limbless. Significant reduction of bones of skull roof.
Discomfort and forebodings: One living group has been neglected: Gymnophiona, the caecilians. Traditionally classified among Amphibia with frogs and salamanders, they are superficially worm-like, limbless, aquatic or burrowing, and possess heavily reinforced skulls even though, like members of Batrachia, their skull bones are fewer in number. The oldest fossil is Eocaecilia from the Early Jurassic - very much a caecilian even though it has paired limbs. Two major phylogenetic hypotheses address their position:
- Monophyletic Lissamphibia: Both Linnean tradition and numerous phylogenetic analyses place them as the sister-taxon to Batrachia in a monophyletic Lissamphibia that unites all of the living amphibians.
- Polyphyletic Lissamphibia: Other plausible analyses place Gymnophiona among Lepospondyli, making them stem amniotes. A recent report potentially connects them with a group of extinct temnospondyls - Stereospondyli (E.G. Mastodonsaurus, with the largest head of any temnospondyl.)
The strange thing: Reptiliomorphs remain primarily aquatic with three exceptions:- Certain microsaurs
- Adult seymouriamorphs (but definitely not larvae)
- Cotylosauria: Containing Amniota and its closest relatives.
Amniota: (Carboniferous - Recent) The most recent common ancestor of mammals and birds and all of its descendants. Amniotes represent a major departure in the evolution of full terrestriality in vertebrates, and are the first vertebrates fully to break the dependance on bodies of water.
Synapomorphies:
Amblyrhynchus cristatus the Galapagos marine iguana- The amniotic egg: An air-breathing egg characterized by a shell and extraembryonic membranes. These enclosed the amniote embryo in a private pond during its development and mediated gas-exchange with the external environment.
- Claws or nails at the end of digits. This novelty allows digging, and burying of eggs. (So are these characters fully independent?)
- An egg-tooth on the snout to facilitate hatching. (Again independent?)
- Complete loss of water-breathing larva.
- Complete loss of lateral line system
- Internal fertilization by means of a penis.
- Two sacral vertebrae.
- Formation of the astragalus (a large tarsal that articulates with the tibia) from the fusion of at least three previous tarsals.
In a way, claiming that these are seven separate characters seems dishonest since all but the last two are associated with one important life-style change: All amniotes reproduce using an air-breathing egg or its derivative.
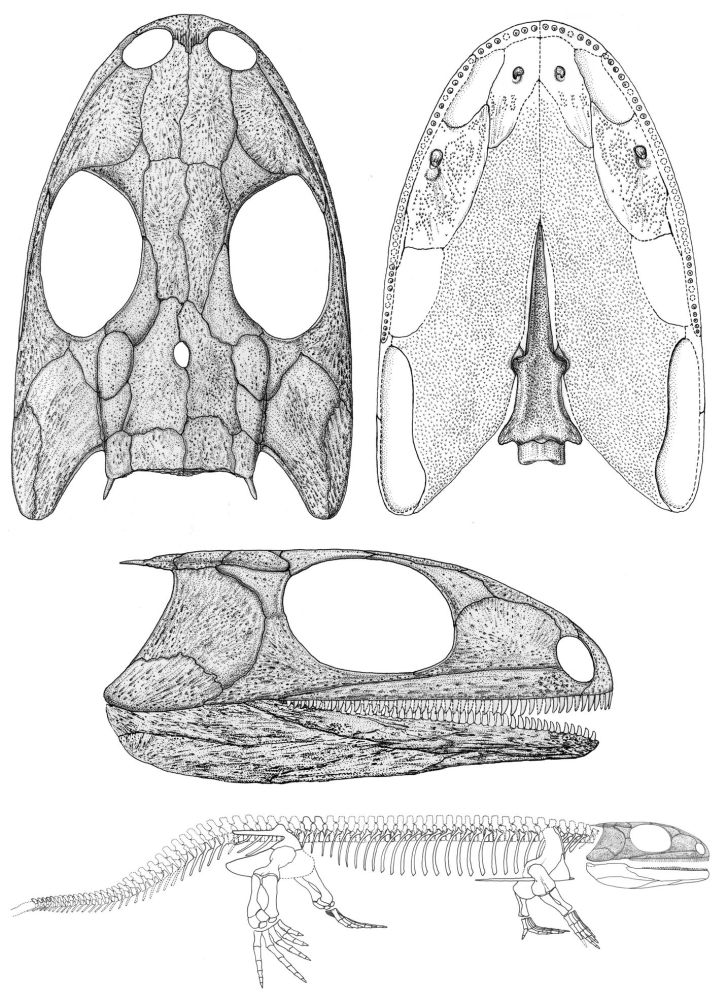
The common ancestor of amniotes is unknown. The earliest fossil amniote, Hylomonus dates from the Late Carboniferous (312 mya). From the Carboniferous and Permian we have fossils of candidates for its sister-taxon, including:- Diadectomorpha (Latest Carboniferous - Early Permian) Including the unspecialized predatory Limnoscelis and Tseajaia, and the the dog-sized Diadectes (right), the first known tetrapod herbivore. These lack claws, however Diadectes and some relatives show fusion of the astragalus as adults but not as juveniles. Aquatic larvae are unknown for them and they represent the first group with large terrestrial adults.
Stupid Amniote Tricks: By eliminating the requirement to return to moist environments to mate, amniotes gained access to dominant terrestrial ecological niches. A detailed account of their diversity and evolution is beyond the scope of this course, but there are some major themes.
Fossil diversity:
- Synapsida: (Carboniferous - Quaternary) Amniotes more closely related to mammals than to birds.
- The most primitive form the paraphyletic group "Pelycosauria" (from Late Carboniferous to Early Permian). Dominated the first terrestrial vertebrate faunas, with:
- Small predators (E.G. Varanodon agilis)
- Large herbivores (E.G. Cotylorhynchus)
- Large predators (E.G. Dimetrodon - right)
- Therapsida: (Permian - Quaternary) Mammals and their close relatives - dominated the faunas of the later Permian and persist into the present as mammals.
- Large herbivores Dicynoodntia (right)
- Large predators Gorgonopsia (right)
- Small therapsids survive the P-Tr extinction, ultimately giving rise to Mammalia in the Jurassic.
- The most primitive form the paraphyletic group "Pelycosauria" (from Late Carboniferous to Early Permian). Dominated the first terrestrial vertebrate faunas, with:
- Sauropsida: (Carboniferous - Quaternary) Amniotes more closely related to birds than to mammals.
- Parareptilia: (Permian - Triassic) Range from small generalized reptiles to large specialized forms. Some highlights:
- The first facultative bipeds (Eudibamus)
- Large herbivorous Pareiasauria
- Small armored Procolophonia (right)
Diapsida: (Carboniferous - Quaternary) Reptiles characterized by elongate limbs in which the astragalus is functionally integrated with the tibia, and the skull is specialized for a rapid bite. Encompasses minor groups (E.G. Drepanosauridae - right) and three major ones:- Lepidosauria: Lizards and rhynchocephalians and their relatives, including snakes and mosasaurs.
- Euryapsida: (Triassic - Cretaceous) The major marine reptiles of the Mesozoic, including:
- Eosauropterygia: (Triassic - Cretaceous) Plesiosaurs and their kin.
- Placodontia: (Triassic) Specialized shell-crushers
- Ichthyopterygia: (Triassic - Cretaceous) Fish-like predators of shallow and deep marine water.
- Archosauria: The dominant land vertebrates of the Mesozoic.
- Crurotarsi: (Triassic - Quaternary) Crocodilians and their fossil relatives, including:
- Large herbivorous Aetosauria (Triassic - right).
- Top predators - "Rauisuchia" - (Triassic).
- Crocodilomorpha: Crocodilians and their fossil relatives
- Ornithodira: (Triassic - Quaternary):
- Pterosauria: (Triassic - Cretaceous): The original flying reptiles (right).
- Dinosauria: (Triassic - Quaternary): Dinosaurs.
To Next Lecture.
To Previous Lecture.
To Syllabus.
- Crurotarsi: (Triassic - Quaternary) Crocodilians and their fossil relatives, including:
- Parareptilia: (Permian - Triassic) Range from small generalized reptiles to large specialized forms. Some highlights:

Dimetrodon by D. Bogdanov. - Vertebral centra: These were small or absent in aquatic sarcopterygians, which retained an exposed notochord. Movement on land demanded that the notochord be strengthened against bending and compression. Thus, robust centra formed as ossifications of the notochord.
- Paired limbs became larger, more robust, and more flexible. Remember, a tristichopterid, whose radius was twice the length of its ulna, could not have bent its arm effectively.












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}