


Key Points:
•Protostomia is divided into two major branches, Ecdysozoa (arthropods and kin) and Spiralia. Within the latter are numerous small-bodied non-fossilizing forms and Lophotrochozoa (brachiopods, bryozoans, annelids, mollusks, and kin.)
•Mollusca is among the most diverse clades of metazoan: second only to Arthropoda in species diversity. The presence of a calcareous shell in most taxa gives them an unparalleled fossil record among animal groups.
•A series of body and trace fossils seem to document stem-mollusks running from the Ediacaran to the Cambrian Explosion.
•Testaria (crown-Mollusca) includes two major divisions: Aculiferans (polyplacophorans and aplacophorans) and the extraordinarily diverse Conchifera ("monoplacophorans", cephalopods, scaphopods, gastropods, bivalves, and kin). Relationships among conchiferan clades remains poorly resolved.
•Cephalopoda is primarily a clade of predatory forms. The chambered shell allows for nektonic movement of various degrees.
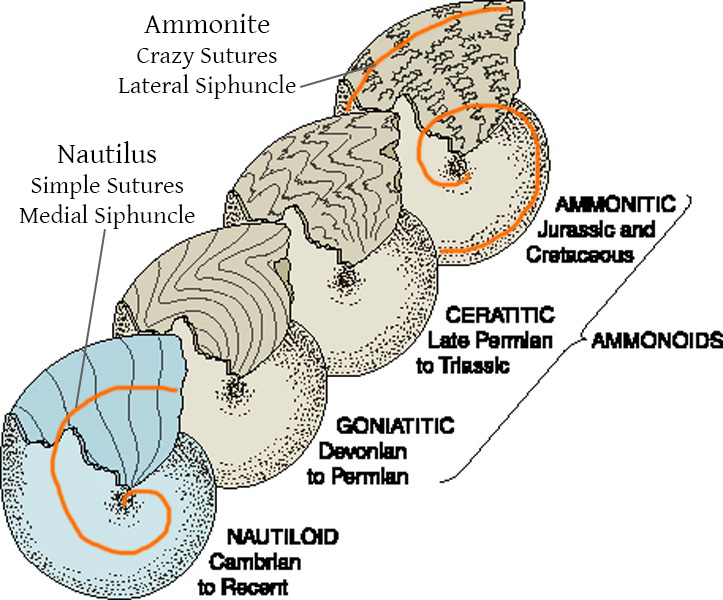
•"Nautiloids" are a paraphyletic assemblage of basal cephalopods. Different clades evolved differing adaptations to deal with the conflict between buoyancy generated within the phragmocone and the center of gravity from the body mass.
•Ammonoidea was a very successful clade of shelled cephalopods of the Devonian through the end of their Cretaceous. At least some members moved away from a predaceous mode of life to planktonivory.
•Coleoidea contains the majority of living species. In this clade the shell was successive internalized, reduced, then lost all together. Given their soft-bodied nature, their fossil record is generally poor outside of Belemnoidea.
"Men may sail the seas for a lifetime and seldom, if ever, come in contact with the nightmare monsters that inhabit the caves and cliffs of the ocean floor. Gazing down at the slightly muddy water, the men of The Unicorn saw a squirming mass of interwoven tentacles resembling enormous snakes, immensely thick and long and tapering at their free ends to the size of a man's thumb. It was a foul sight, an obscene growth from the dark places of the world, where incessant hunger is the driving force. At one place, down near the bulge of the hull, appeared a staring gorgon face with great lidless eyes and a huge parrot beak that moved slightly, opening and shutting as though it had just crunched and swallowed a meal of warm flesh." -- "Fire in the Galley Stove", William Outerson (1937)
PROTOSTOMIA: DIVISIONS
We have seen that Bilateria is subdivided into to major divisions, Protostomia and Deuterostomia. We will explore the diversity of the latter later in this course. For now, it is time for the Protostomia.
Take a look again at this simplified potential cladogram of metazoans:
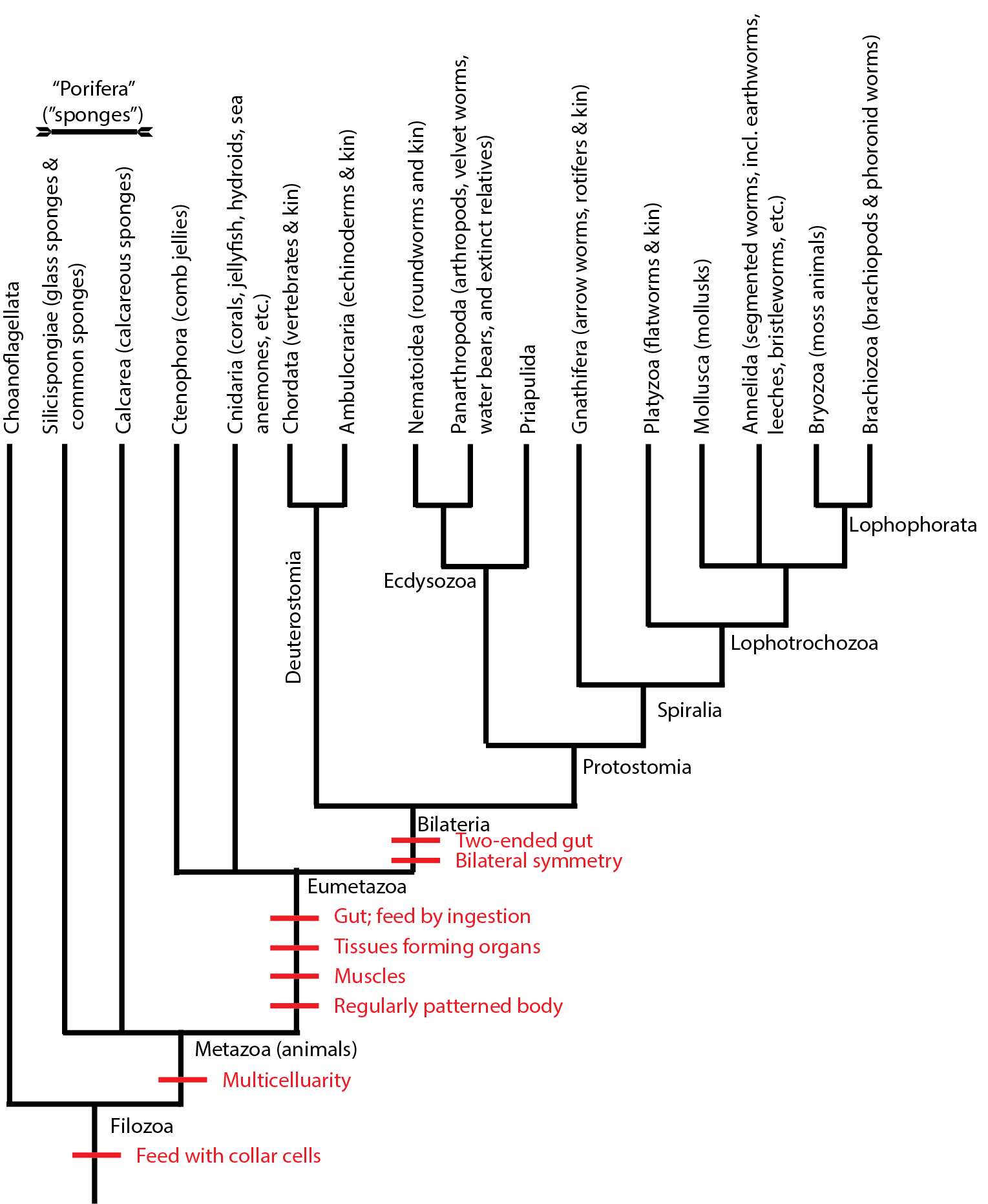
Protostomia seems to divide into two clades. One of these is Ecdysozoa, the "moulting animals". The prominent member of this clade is Panarthropoda (arthropods and kin), as well as nematodes, priapulids, and others.
The sister group to Ecdysozoa is Spiralia, named after the spiral cleavage in protostomes. Arrangements within Spiralia remain poorly resolved. In many studies the basal branches of Spiralia are micro- and meiofauna taxa with almost no fossil record: the Gnathifera (chaetognaths, rotifers, and their kin) and the hugely diverse (and one with many fossilizing members) Lophotrochozoa. This is so-called as it contains the (probably) paraphyletic "Trochozoa" as well as the (possibly) monophyletic Lophophorata (Bryozoa, Brachiopoda, Phoronida, Entoprocta). See the previous lecture on lophophorates.
The "Trochozoa" are noted for having a trochophore larval stage (with a wheel-like band of cilia around them. The predominant members of "Trochozoa", however, are Annelida (the segmented worms) and Mollusca (the mollusks).
We'll briefly examine annelids here. Living annelids include the (paraphyletic) marine polychaetes ("bristle worms"), the (paraphyletic) terrestrial oligochaetes (earthworms), the (monophyletic, and nested within oligochaetes) Hirudinea (leeches), and a smattering of micro- and meiofaunal clades scattered amonst the rest of these. It's a wormy mess.
Polychaetes represent the basal form in annelids, and not surprising there is a high diversity of such form in Cambrian Lagerstätten. A notable fossil variety is the Ordovician-Carboniferous Machaeridia. This clade has calcitic dorsal sclerites: if found isolated and in the earlier Cambrian, these elements would easily be considered members of the "Small Shelly Fauna".
Now, onto Mollusca, the "soft ones". Among animal clades they are second only to Arthropoda in diversity, and the fact that most groups have macroscopic calcareous shells gives them both an phenomenal fossil record as well as a huge modern amateur following of "conchologists" (in addition to professional malacologists).
The fossil record of crown-group mollusks extends back into the Cambrian, but the stem of mollusks seems to begin in the Ediacaran. Below is a possible phylogeny of Mollusca:
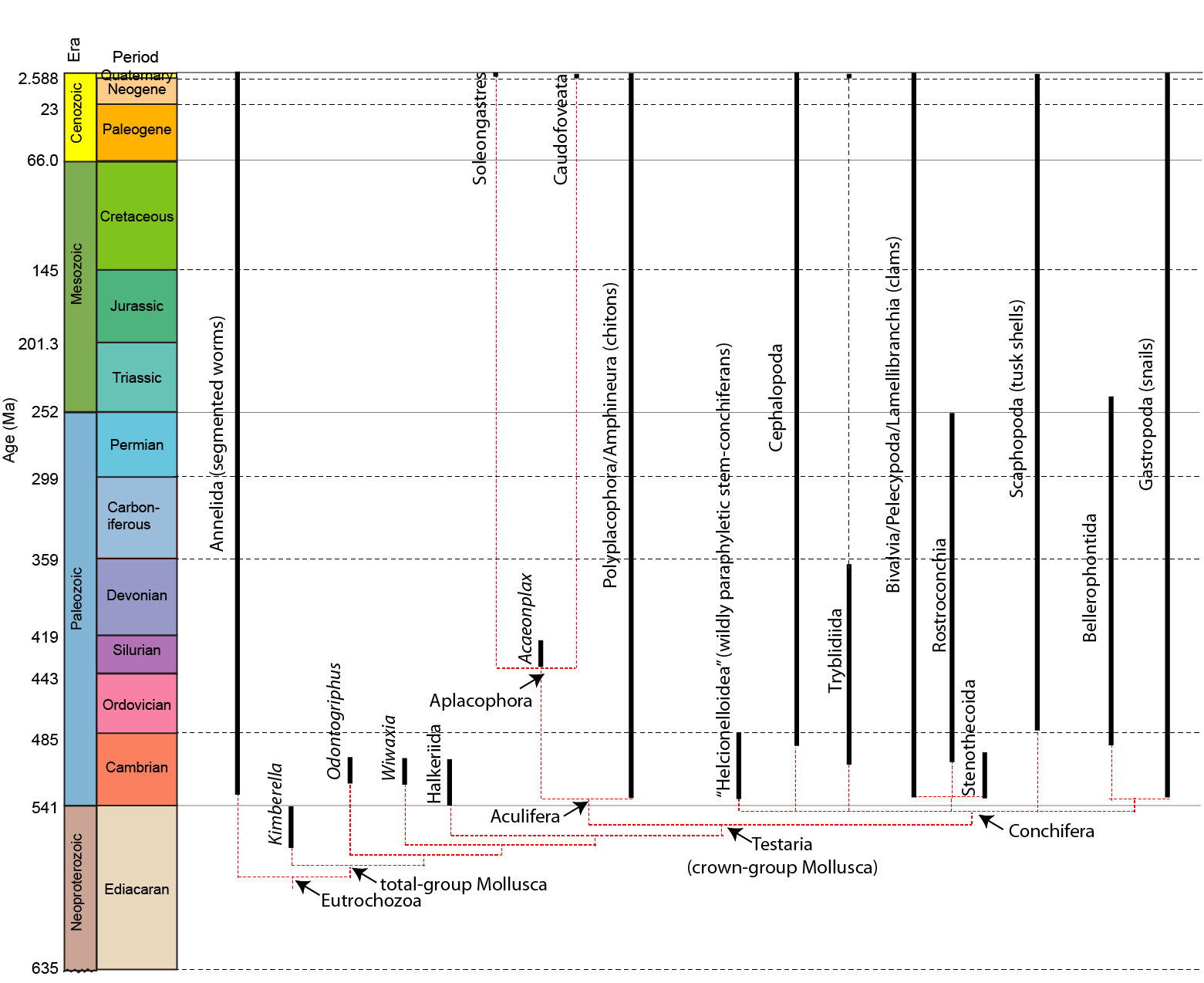
The exact definition of "Mollusca" isn't agreed upon: the crown-mollusk group is Testaria. But if primitive Ediacaran Kimberella is a basal mollusk, then the following traits are inferred at the base of the clade:
Given the lack of internal body anatomy, it is uncertain if other soft tissue molluscan traits were present, such as:
Cambrian near-shore sandy environments can contain the ichnite Climactichnites, which seems to the belly foot trace of a very large mollusk (up to about 10 cm wide!). There are no shells of giant conch-sized mollusks at this time, so the Climactichnites maker was either shell-less or had a sclerotome (skeleton) made of individual tiny sclerites.
Odontogriphus of the mid-Cambrian Burgess Shale is a bilaterian with a belly foot, a fringe of gills, a domed top, and a mouth with a radula: almost certainly an early stem-mollusk.
Wiwaxia is a small domed Burgess Shale bilaterian covered in tiny chitinous plates and spines. Initially considered a short domed stem-annelid, the discovery it had a radula makes it more reasonably a stem-mollusk. One question, though, is the relation of the chitinous plates to chaetae of polychaete annelids and/or the sclerites of the next group.
Halkeria and allies in the Halkeriida of the earlier-through-mid-Cambrian resemble enlarged, elongate Wiwaxia, but where the plates have become calcitic sclerites. Some of these individual sclerites closely resemble Small Shelly Fauna fossils, and also some of the plates of aplacophorans (about which see below). This phase is quite reasonably close to being an outgroup to crown-mollusks.
Speaking of which, the crown-members of Mollusca form the clade Testaria (the "shelled ones"), united by following synapomorphies:
There are seven major extant clades of mollusk:
Relationships among Testaria are problematic but resolving... Nearly every molecular and many morphological studies support a union of aplacophorans and polyplacophorans into Aculifera. The remaining molluscs form the clade Conchifera, ancestrally sharing a single slightly conical shell. Disregarding for a moment the living and extinct "monoplacophoran" grade, traditional mollusk systematics united gastropods, scaphopods, and cephalopods into Cyrtosoma (more encephalized; shell taller and more conical), with bivalves and some extinct clades as Diasoma (shells dorsoventrally elongated). Other analyses put scaphopods with bivalves, united by a protrusible foot used in burrowing. In contrast, recent molecular studies unite Gastropoda, Scaphopoda, and Bivalvia into Pleistomollusca (which thus contains 95% of extant mollusk species!!), and leaving cephalopods as either the sister group to tryblidiids or as the sister group to pleistomollusks.
Lacking any significant resolution, we'll leave the conchiferans as an unresolved polytomy.
Polyplacophora (chitons) (also called Amphineura)
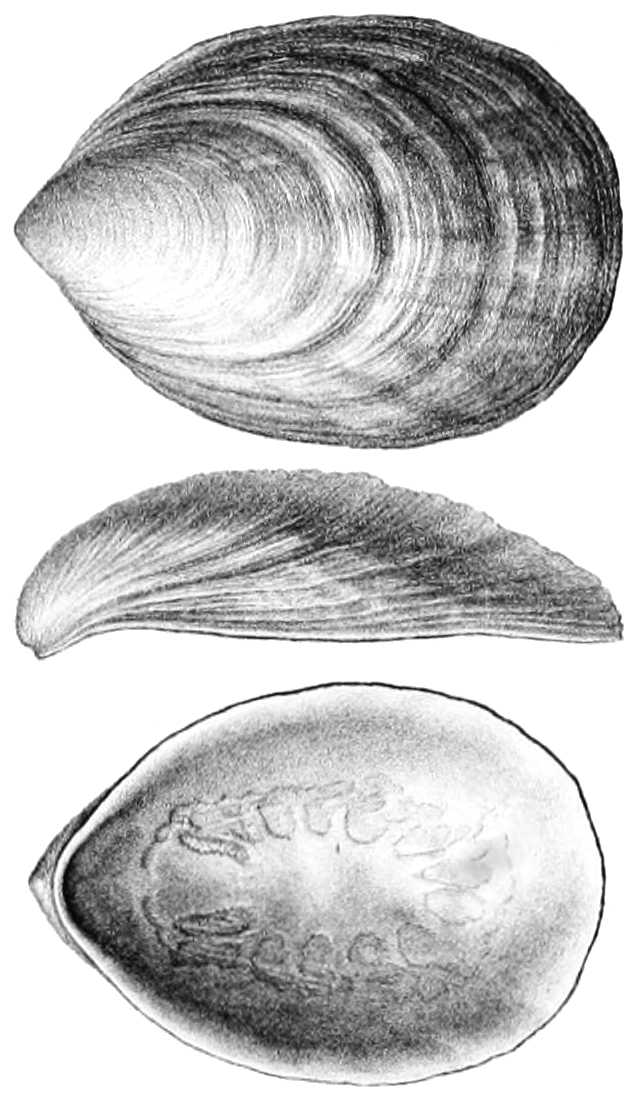
The term "Monoplacophora" was proposed in 1940 for a set of early and mid-Paleozoic univalve mollusks previously interpreted as limpet-like gastropods. Unlike gastropods these shells had multiple pairs of muscle scars along the interior surface. This indicated that the internal meat was NOT torted.
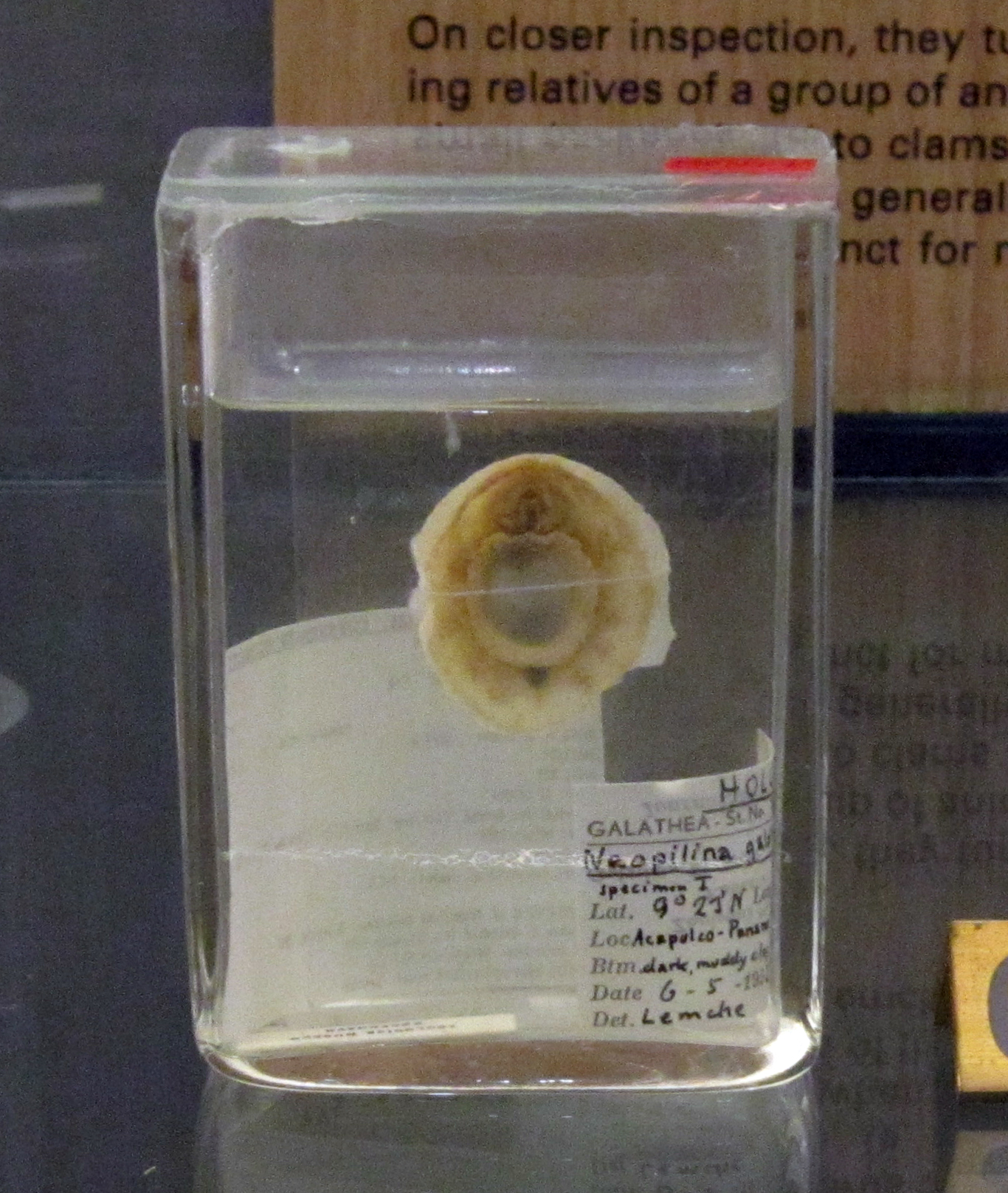
In a tremendous surprise, in 1952 a living monoplacophoran was discovered off Costa Rica! Subsequently 30 additional living species have been found.
Living monoplacophorans have:
The living monoplacophorans are part of a clade Tryblidiida. However, the univalved untorted epifaunal morphology is simply the basal condition of the clade Conchifera. So "monoplacophorans" are simply a paraphyletic grade of any conchiferan not lucky enough to have evolved into one of the recognizable subclades.
Another notable "monoplacophoran" clade was the Cambrian Helcionelloida. These are higher-spired shells than in tryblidiids, and have an exhalent pipe presumably used in respiration. These are part of the SSF.
Some other "monoplacophoran"-grade taxa will be discussed in relation to their more derived relatives.
A phylogeny of the cephalopods:
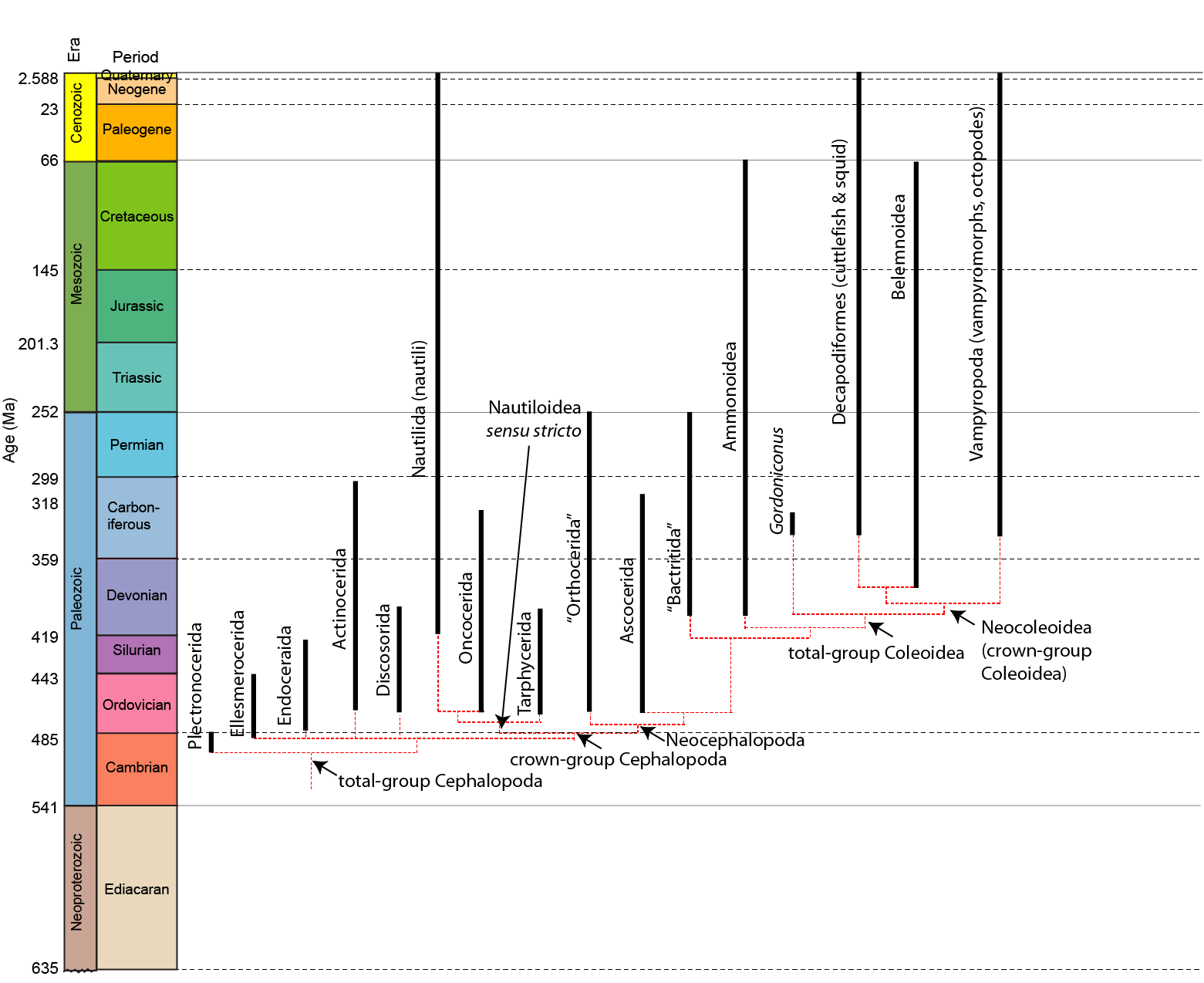
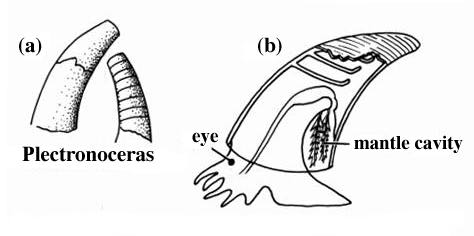
Basalmost form: Plectronoceras of Upper Cambrian
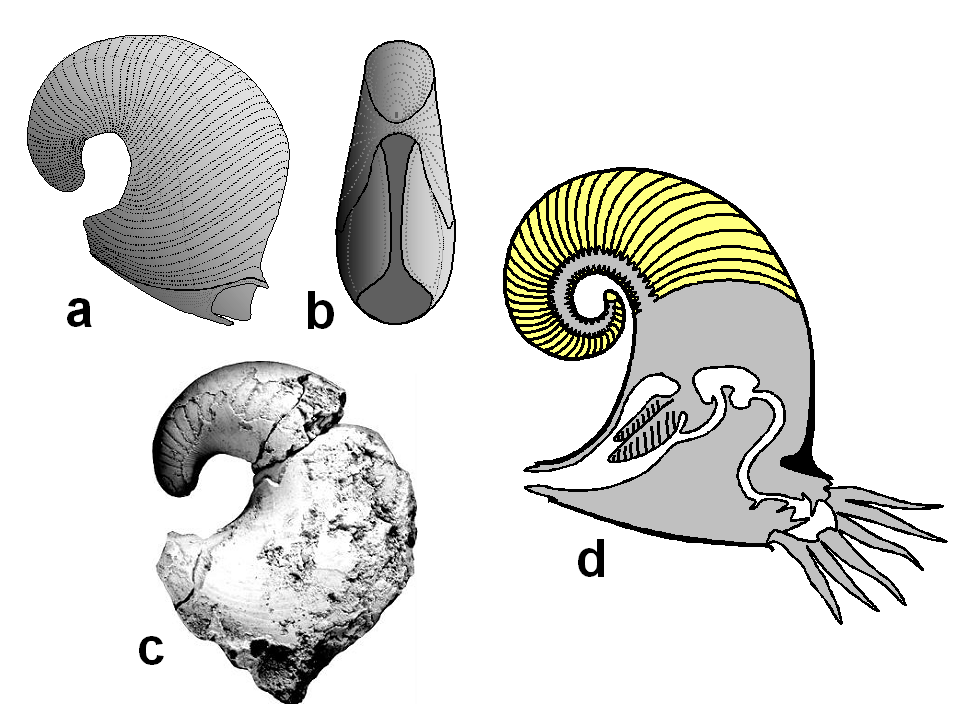
The basal shelled cephalopods have been collectively called "nautiloids", a paraphyletic grade. Ranging from the Ordovician to today, "nautiloids" have rather simple suture lines.
Ancestrally orthocone, cephalopod shells would have been vertically oriented (something nearly EVERY life restoration gets wrong!) due to the center of buoyancy compared to the center of gravity. Different clades attempted to "lateralize" or "horizontalize" the body through different means: for instance, some have cameral deposits (addition of calcareous material into the camerae to weigh-down the ventral size). Additional at least some form have ectochlear (external-to-shell) tissues
Some of the notable "nautiloid" groups: Endocerida
Actinocerida
Discosorida
Nautilida
Orthocerida
Ascocerida
To Syllabus.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}