Key Points:
•The formation is the fundamental unit of stratigraphy.
•The Principle of Faunal Succession facilitates the correlation of rock units across wide distances.
•Biostratigraphy depends on index fossils.
•Biostratigraphic units are rock units, not time units.
•Biozones are defined based on first and last occurrence data.
•Biases of the rock record and quirks of evolution impart difficulties to the interpretation of biostratigraphy.
•Biochrons are time units.
•Quantitative biostratigraphy allows fossils to address questions about sedimentation and depositional processes.
"Organized Fossils are to the naturalist as coins to the antiquary; they are the antiquities of the earth; and very distinctly show its gradual regular formation, with the various changes inhabitants in the watery element.
(William Smith, 1817. Stratigraphic System or Organized Fossils.)
Stratigraphy without fossils:
Paleontology can illuminate important issues in both biology and geology. Today's topic addresses one of paleontology's greatest applications in the service of geology.
Stratigraphy: The detailed history of Earth's surface revealed by the order and relative position of superposed rock units.
Using the stratigraphic principles of Nicholas Steno: (1668) and the uniformitarian principle of James Hutton (1795), Geologists of the early 19th century could establish the relative ages of rock units on the formation scale.
Contact of the Coconino (above) and Hermit Shale formations - Grand Canyon
Formations: the smallest mappable rock units. E.G. the Chinle Formation. Remember, formations must:
Have a distinct lower and upper boundary
Have a type section: a particular spot where it is well exposed and can be compared to other localities
Have been laterally continuous during formation (even if its surface exposure isn't)
Be stratigraphically continuous from top to bottom (except for intertonguing)
Be mappable on a standard quadrangle-scale map
Coconino Formation
Typically have a characteristic range of lithologies. E.G. the Permian Coconino Formation of northern Arizona. This mature eolian quartzarenite is exposed both in the Grand Canyon (right) and in nearby Walnut Canyon, and is continuous between them beneath the surface.
Although any kind of rock can provide useful information, stratigraphers are typically interested in sedimentary rocks (sandstone, limestone, shale, etc.) formed from the transported remains of preexisting rock.
Give us Time Lines!
:
If we want a global history of Earth's depositional environments, we really want to identify time lines in the rock record: geographically widespread surfaces that formed synchronously as a result of the same events, like:
In the early 19th century, looking strictly at the rocks, themselves, these were not easy to find, and stratigraphers had to address confusing aspects of the rock record like:
Physical proximity of formations: Units had to be seen in direct association for use to work out their relative ages.
Diachronous (i.e. Time transgressive) units representing a particular depositional environment might overlap temporally with adjacent units causing confusion over whether superposition of rock units actually signifies relative age.
But as it happens, many fossils provide decent, widespread approximations of time-lines.
Index Fossils, Correlation, and the birth of Biostratigraphy
In 1796 William Smith, a British civil engineer, addressed this, adding the principle of Faunal succession to those of Steno and Hutton. Essentially, Smith noted that:
Rock units were characterized by unique sets of fossil taxa.
These sets of fossil taxa - faunas presumably represent the diversity of things living at the time the sediment was laid down.
That the occurrence of many fossils was independent of the lithology of the rock.
In essence, changes in fossil faunas function as stratigraphic time lines.
By noting the fossils present, it became possible to:
correlate rock units of varying lithologies across vast distances
establish time horizons in lithologically uniform or diachronous rock units.
In 1815, Smith published the first geologic map of England and Wales (or anything - right).
The great thing: These correlations made the association of formation scale units into larger ones (the stages and systems on which the Geologic Time Scale is based) possible. Thus, essentially all stratigraphy above the formation scale depends on biostratigraphy.
The ammonite Sphenodiscus, restricted to Maastrichthian Stage of Cretaceous.
Which fossils do we use?
Index fossils: Fossils useful in biostratigraphy.
Common
Geographically widespread
Easily preservable
Diagnosable
Found in multiple environments (when dead)
Short species duration
Example: Ammonoids (right), Shelled cephalopods that evolved quickly, so each species lasted only a few million years, but whose remains were distributed worldwide in many environments.
Facies fossils: In sharp contrast. Fossils of organisms that endured for long periods of geologic time but were linked to a specific environment. Example: Lingula (right), a brachiopod living only in lagoonal mud-flats that has changed very little in the last 500 million years.
Rock units are not time units!
With Steno's and Smith's principles as a basis, geologists define a heirarchy of higher order rock units, including:
Stages: Groups of formations characterized by distinct faunal assemblages. E.G. the Carnian Stage of the Upper Triassic Series.
Series: Groups of stages. E.G. the Upper Triassic Series
Systems: Groups of series. E.G. the Triassic System
Larger units need not be contiguous in space but are assumed to be contiguous in time. Their upper and lower boundaries must be instantaneous and isochronous. From these, we derive the Geologic Time scale, in which geochronologic Periods correspond to lithostratigraphic Systems.
Remember, when we talk about a formation, stage, series, or system, we are talking about a bunch of rock, not an interval of time. The numerical dates that we place on their upper and lower boundaries are secondary to the identity of the rock units. For example, the formal base of the Jurassic is a stratotype - a specific contact in the Kuhjoch section of the Karwendel mountains in Austria, between rock units dated to 201.3 Ma. If we were to discover that Triassic rocks below that contact were actually only 200 Ma, we would not say, "Oops. I guess the rocks of the stratotype are Jurassic after all." Rather, we would revise the age of the boundary between the Triassic and Jurassic.
Subsequent to Smith:
1842 - Alcide d'Orbigny (1802-1857) published analysis of Jurassic system of France, and discoverer of foranimiferans. He established the concept of stages which he supported by identifying fossil assemblages. Several stage names still in use were coined by him. A 19th century naturalist with one foot in the future and the other in the past. He regarded each stage as an interval between 27 separate supernaturally mediated creations and flood-induced extinctions.
Roughly contemporaneously, Friedrich Quenstedt (1809-1889), a mineralogist and paleontologist, established his own system based on detailed data-bases that introduced the use of first and last appearance data of individual species, which he considered more precise than d'Orbigny's assemblages, which he viewed as too vague.
Quendstedt's student Albert Oppel (1831-1865) studied biostratigraphy of France, Switzerland, and England, augmenting Quendstedt's system and developing a system of dignostic aggregates identified by overlapping range zones called Biozones that marry d'Orbigny's assemblages with Quenstedt's use of first and last occurrence data. By achieving this, Oppel is widely viewed as the founder of modern biostratigraphy. Coined the term "index fossil."
So let's learn more about bio zones.
Biozones - Rock units!
Primary data of biostratigraphy: presence or absence of a fossil taxon in a geologic horizon
Last Appearance Datum (LAD): either local or global
First Appearance Datum (FAD): either local or global
Biozone (often just "zone"): Rock unit characterized by one or more taxa that permit it to be distinguished from adjacent rocks.
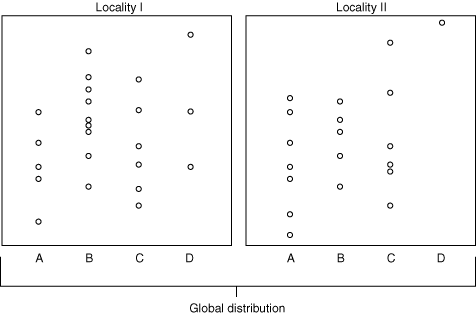
The following examples are drawn from the hypothetical microcosm at right. As we explore these, note that the definition of most biozones requires some element of uncertainly or inferrence.
Types of Biostratigraphic units (and thus rock units):
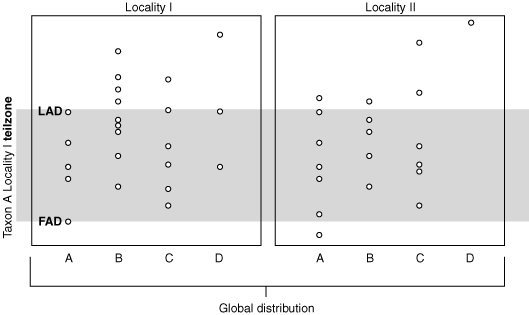
Teilzone of taxon A at locality I.
Teilzone: Between local FAD & LAD of that taxon. - The truly unambiguous observations!
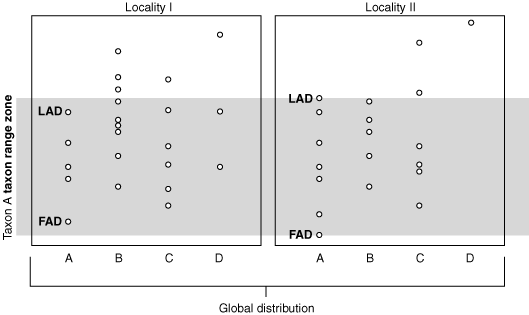
Taxon Range Zone of taxon A
Taxon Range Zone: Between global FAD & LAD of that taxon. - Requires some inferrence - Global correlation of horizons bearing the fossil taxon.
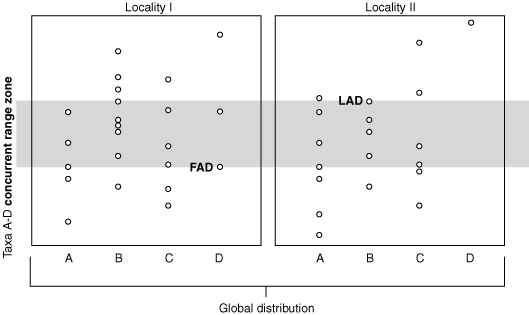
Concurrent Range Zone of taxa A - D
Concurrent Range Zone: Intersection of the taxon range zones of two or more taxa.
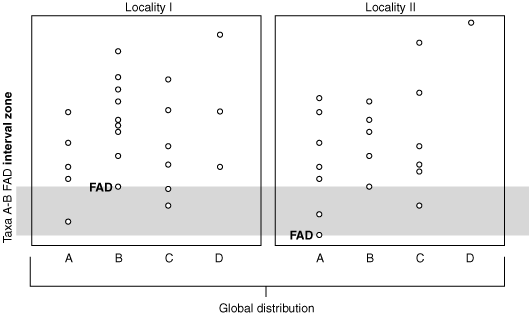
Interval Zone of taxon A and B FADs
Interval Zone: Interval (global) between two successive FADs or two successive LADs
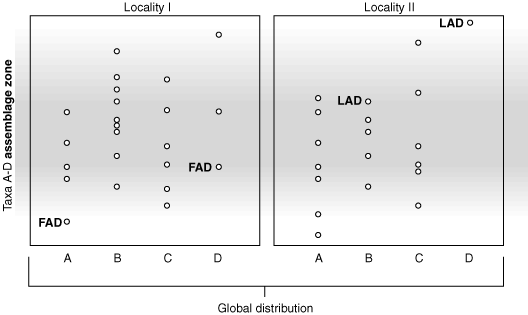
Assemblage Zone of taxa A - D
Assemblage Zone: Characterized by 3 or more taxa in natural assemblage. Biostratigraphers like assemblages because they are free from some of the complications that plague biozones based on a single taxon. (See next lecture) Alas, with assemblages, they must put up with fuzzy boundaries, as the FADs and LADs of the constituent taxa aren't simultaneous.
Special Case: Oppel Zone: Addresses the shortcomings of the assemblage zone by being defined by the FAD or LAD of one taxon, but characterized by additional taxa of an assemblage. Named for Albert Oppel, the first to use non-arbitrarily defined biozones (1858).
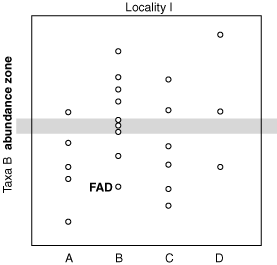
Abundance Zone of taxon B
Abundance Zone (also called Peak Zone, Acme Zone): Subset of teilzone where index species reaches some higher level of abundance: useful locally, but almost certainly environmental rather than time-related. That isn't to say that there is no time signal necessarily - Note examples of global changes in abundance due to global environmental changes. E.g.:
The coccolithophorid Braarudosphaera which bloomed in times of global environmental stress.
Changes in coiling direction of the foram Globotruncana truncatulinoides in response to global temperature changes.
Reasons for caution
Biostratigraphy opened the door to global correlation of strata but is, nevertheless, subject to biases and filters that make it more reliable on a local scale than a global one.
Biases of the Rock Record
This never happened!
There are no perfect index fossils.
Even good ones are subject to some substrate/facies constraints. Thus, all fossils are, to some degree, facies fossils. An ammonoid, for example might have a global marine distribution, but will still never turn up in terrestrial deposits.
Species' geographic ranges change over time. Thus, many critters' records contain some biogeographic signal, such that local FADs and LADs may record immigration and extirpation. In effect, critters, as well as rock units, can be diachronous.
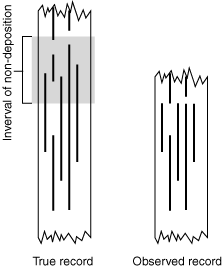
The rock record is incomplete and no rock unit preserves ancient environments on a global scale. Thus:
Abrupt "special-creation-like" appearances of evolutionarily derived creatures. E.G. the placodont Henodus (right). (Compare to a normal placodont.)
Lesson - Evolution (without which biostratigraphy would be impossible) can be difficult to interpret.
Mass Extinctions: Abrupt simultaneous extinctions of numerous taxa, if real, are powerful biostratigaphic markers and are interesting ecological phenomena in their own right. Alas, identifying them isn't straightforward. It requires continuous deposition and a reasonably dense fossil record.
Unconformities create the impression of an abrupt extinction event when in truth, a gradual turnover is occurring. Unconformities are more common than true mass extinctions. Consequently, they are the most likely cause of abrupt simultaneous disappearances.
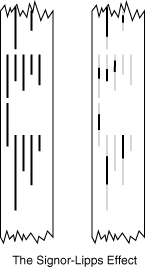
A sparse sample introduces statistical uncertainty into an otherwise good depositional record. Consider the Signor-Lipps effect in which a simultaneous mass extinction is made to appear gradual by random sampling from a poor record.
Confidence intervals: Until now we have concentrated on FADs and LADs, but actually every horizon in which a given taxon occurs is a datum that can be used to constrain its confidence interval statistically. A very sparse record yields wide 95% confidence intervals above and below observed FADs and LADs. A dense record yields narrow 95% confidence intervals. In a case of a single occurrance, the confidence interval is infinite.
Lazarus Taxa: Taxa that temporarily "disappear" and then reappear in fossil record. This might be because of environmental changes, or local extirpation and re-immigration. E.g. North American horses, coelacanths (right).
Miocene tiger shark fossil in Holocene sediments from Polar Trec
Zombie effect: Post-extinction reworking of specimens into younger sediments (right). As in Fasstt et al., 2000, these can be significantly misleading.
Bias of Evolution
Flabellum, a modern scleractinian coral from Wikipedia
Elvis Taxa: Taxa that converge on extinct forms, giving false impression of Lazarus taxa. (A particularly common problem with planktonic forams, whose morphology is strongly biomechanically constrained. Also reef forming organisms, consider Cambrian archeocyathid sponges, late Paleozoic rugose corals and Cretaceous rudistid clams.)
In a biological sense, FADs are arbitrary. Assuming that evolution is gradual, (no consensus on that!) how do we identify the first occurrence of a new species? Indeed, how do we distinguish an evolving lineage from a branching phylogeny (right)?
We can't reliably distinguish true global FADs and immigrations.
Migrating species are highly time-transgressive. Hopefully, we can see that the immigration of a taxon into a region is more nearly instantaneous than a taxon's evolution in place.
Alas, we are constrained by the chronological resolution of the rock record, which is usually too poor for us to distinguish. For example, the Miocene Hipparion event - the immigration into western Eurasia of three-toed horses ended up being two widely separate events
Eofallotaspis, a Cambrian index taxon
The most useful index taxa, although varying with geologic time, have certain characteristics that give them global, but not absolutely universal applicability. Absolutely no group lives in every possible Earth environment across the Phanerozoic. Across time, our reliance must shift:
Cenozoic: planktonic microorganisms, especially foraminiferans
Modern comparisons of biostratigraphy with abiotic time line criteria such as magnetostratigraphy or stable isotope ratios suggest that planktonic organisms in the marine realm, at least, are reasonably reliable.
Species may be time transgressive but assemblages are typically not. In fact, the co-occurance of different taxa is strongly controlled by climatic factors and geographic factors that change over broad regions. E.G. discordant faunas of the Pleistocene. In the modern world, the ranges of:
Barely overlap or don't overlap at all. Phillips, 2006, however, showed that during the last Pleistocene glacial, their ranges overlapped in northern Alabama and Mississippi. Thus, changing temperature was only one environmental factor explaining changing species ranges with the last deglaciation, and the Pleistocene glacial maximum and the Holocene present regional faunas that are distinct from one another, even when the species that make them up are the same.
Biostratigraphic Conventions:
Mindful of these considerations, we see why biostratigraphers employ a variety of zone definitions despite their invocation of conjecture and assumptions: In many circumstances, the ability to bring lots of data to bear on a problem is simply more important than the avoidance of the fuzziness that follows from inference and conjecture. The biostratigrapher seeks the optimal tradeoff for the specific situation.
Lower boundary of Turonian Stage of Cretaceous: Global Standard Stratotype and Point (GSSP)
from KQED
Biostratigraphic nomenclature and golden spikes:
Biostratigraphy is the principal determinant of such important things as period boundaries. Boundaries between periods are arbitrarily decided, but usually involve biostratigraphic markers. Some conventions:
Typically, they are based on the first appearance of a diagnostic taxon but not the last - i.e. they are "topless" boundaries.
The boundary is identified with reference to an agreed upon location somewhere in the world called the Global Standard Stratotype and Point (GSSP.) This is physically marked by a spike driven into the rock. (Referred to as "golden spike," but not really gold. Sorry, treasure-hunters.)
E.G. the Silurian-Devonian boundary (right) was decided to be the base of the Monograptus uniformis (a graptolite) zone. Over forty such golden spikes have been established.
But, how does this help us with the stratigraphy of environments in which fossil preservation is spotty and intermittent?
Biochronology - Time Units:
When biostratigraphic data is combined with numeric age information we can use biozones as the basis for
biochrons - time units! (as opposed to biozones which are rock units)).
Central Alaska during the Rancholabrean NALMA from Geo Currents
One famous application: Land Vertebrate Ages. Originally developed during the early and mid-20th century as North American Land Mammal Ages (NALMAs), these were based on the recognition of the regular co-occurrance of distinct mammalian assemblages. NALMAs answered the need for some kind of biochronological tool for North American Cenozoic terrestrial rock units, but were limited by the fact that Cenozoic mammal fossils tended to occur in isolated pockets, often in places with little precise stratigraphic context. (Cave deposits, for example.) As a result, they could not be used as the basis for proper rigorous biozones based on the rocks in which they were found. Instead, they are tied to the time intervals in which the assemblages are inferred to have lived. For regions like the continental deposits of North America, however, NALMAs are much better than nothing, and more frequently mentioned than proper biozones. The NALMAs of the Pleistocene Epoch, for example are good examples and often cited. E.G.:
Rancholabrean: Named after the Rancho La Brea locality in Los Angeles, CA, characterized by the presence of Bison in association with characteristic Pleistocene forms like Mammuthus. 0.024 - 0.011 ma.
Over a century of development, competing criteria were used in definitions of ages. Today, biostratigraphers must:
Formally resolve contradictions that arise as new information becomes available. E.G: The Chadronian was originally defined by the last appearance of titanotheres and the top of the Chadron formation. Alas, titanotheres are now known from above the Chadronian.
Address the circular reasoning that can result from not being able to tie the ages to well defined stratotypes. (One 1998 study linked a Early Triassic vertebrate biochron with the presence of a primitive ichthyosaur thought to characterize the biochron, however the rocks containing the ichthyosaur had only been deemed Early Triassic because of the presence of a primitive ichthyosaur.)
Nevertheless, as a quick-and-dirty, NALMAs/Land Vertebrate Ages are sufficiently useful that they have been expanded globally and pushed confidently into the Late Cretaceous as Land Vertebrate Ages. But note: The use of Land Vertebrate Ages is a tradition among North American paleontologists and stratigraphers originally born of necessity but now, perhaps, just a tradition. Prothero, 1995 reviews the application of new data from improved radiometric dating and magnetostratigraphy, and concludes that the time is ripe for incorporation of NALMAs into proper biozone-based biostratigraphy.
All rest on the assumption that biostratigraphic units are good proxies for time. As indicated above, this seems to be a good first-order approximation.
Final thoughts
Whatever their limitations, biozones are very useful stratigraphic and chronological markers.
Unlike radiometric dating methods, biozones don't lose precision or resolution with increasing age. In this way, they resemble magnetostratigraphic or chemostratigraphic zones.
Can be used in conjunction with other dating techniques. See magnetobiochronology.
Can provide quantitative information about rates of deposition using the methods of quantitative biostratigraphy.
Quantitative Biostratigraphy
Besides hopefully constraining their age and sequence, does biostratigraphy add to our kowledge of the deposition of sediments? Actually, yes.
Graphic correlation: method for stratigraphic correlation based on statistical correlation of first and last appearances, but not biozone terminology. Facilitates comparison of locality sections containing local FADs and LADs of the same taxa. Used to:
Identify errors and outliers.Consider the following data:
Taxon
Section X FAD
Section X LAD
Section Y FAD
Section Y LAD
A
0
3
0
2
B
0
6
0
4
C
1.5
12
1
6
D
4.5
6
3
8
E
4.5
10.5
3
7
F
7.5
10.5
5
7
G
9.75
13.5
6.5
9
H
10.5
15
7
10
I
10.5
15
8
10
J
12
15
8
10
Characterize differences in depositional rate: Consider the following data:
Taxon
Section X FAD
Section X LAD
Section Y FAD
Section Y LAD
A
0
1
0
2
B
1
3
2
6
C
3
4
6
8
D
5
6
10
12
E
5
8
10
13
F
7
9
12.5
13.5
G
10
13
14
15.5
H
11
12
14.5
15
I
12
14
15
16
J
14
18
16
18
Identify depositional hiatuses. Consider the following data:
Taxon
Section X FAD
Section X LAD
Section Y FAD
Section Y LAD
A
0
2
0
2
B
1
4
1
4
C
2
5
2
7
D
3
5
3
8
E
4
5
4
9
F
5
6
7
10
G
5
9
8
12
H
9
11
I
10
14
12
14
J
13
16
13
18
Composite standards: The examples above correlate teilzones from pairs of localities. On a larger scale, the data used to achieve this can be combined into substantial composite standard databases that:
Represent the summation of large volumes of regional data. (Thus, they are neither teilzones nor taxon range zones.)
can also incorporate numerical age data from reliable marker beds.
Comparison of information from a particular locality with the composite standard facilitates robust identification of range extensions, hiatuses, and outliers.
Moral:
No method of biostratigraphy lacks significant biases and limitations, however biostratigraphy compensates by enabling stratigraphers to bring an overwhelming quantity of data to bear, making it cumulatively very powerful.

























{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}