


Key Points:
•Chordata, sister taxon of Ambulacraria, contains Cephalochordata, Urochordata, and Vertebrata.
•Characterized by notochord, hollow dorsal nerve cord, endostyle, myomeres, and post-anal tail.
•Pikaia of the Burgess Shale is just outside of Chordaata.
•Cephalochordata comprises suspension-feeders employing their large pharynx, but capable of fish-like locomotion.
•Urochordata comprises creatures that, as adults, are mostly pharynx, but as larvae display the synapomorphies of Chordata.
•Vertebrata comprises chordates with heads that contain brains and three pairs of special sense organs.
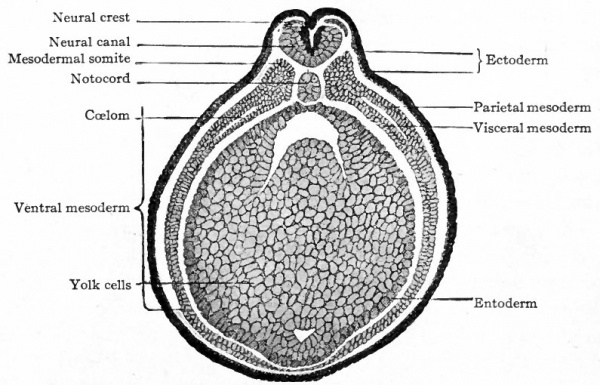
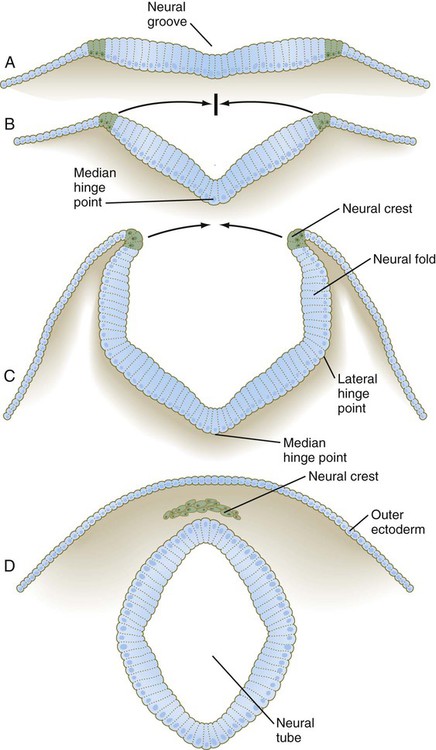
•Developmentally, chordates add the formation of the neural tube to the sequence discussed earlier for Bilateria.
•Vertebrates add to this the formation of neural crest tissue - arguably a consequence of the duplication of HOX genes.
•Urochordata is the sister taxon of Vertebrata, forming Olfactores.
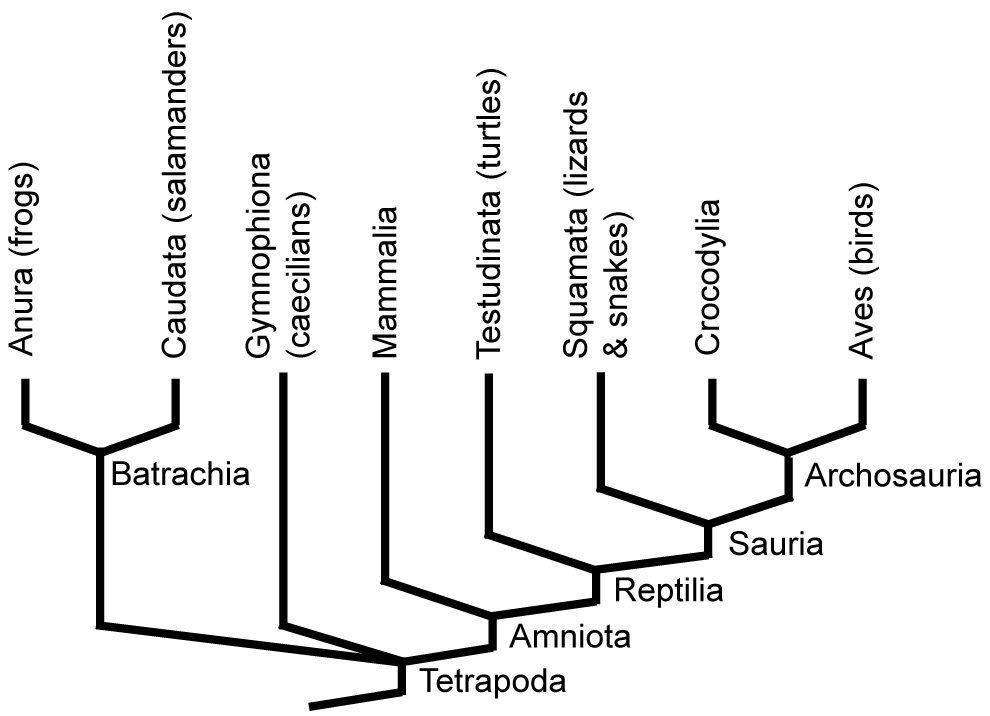
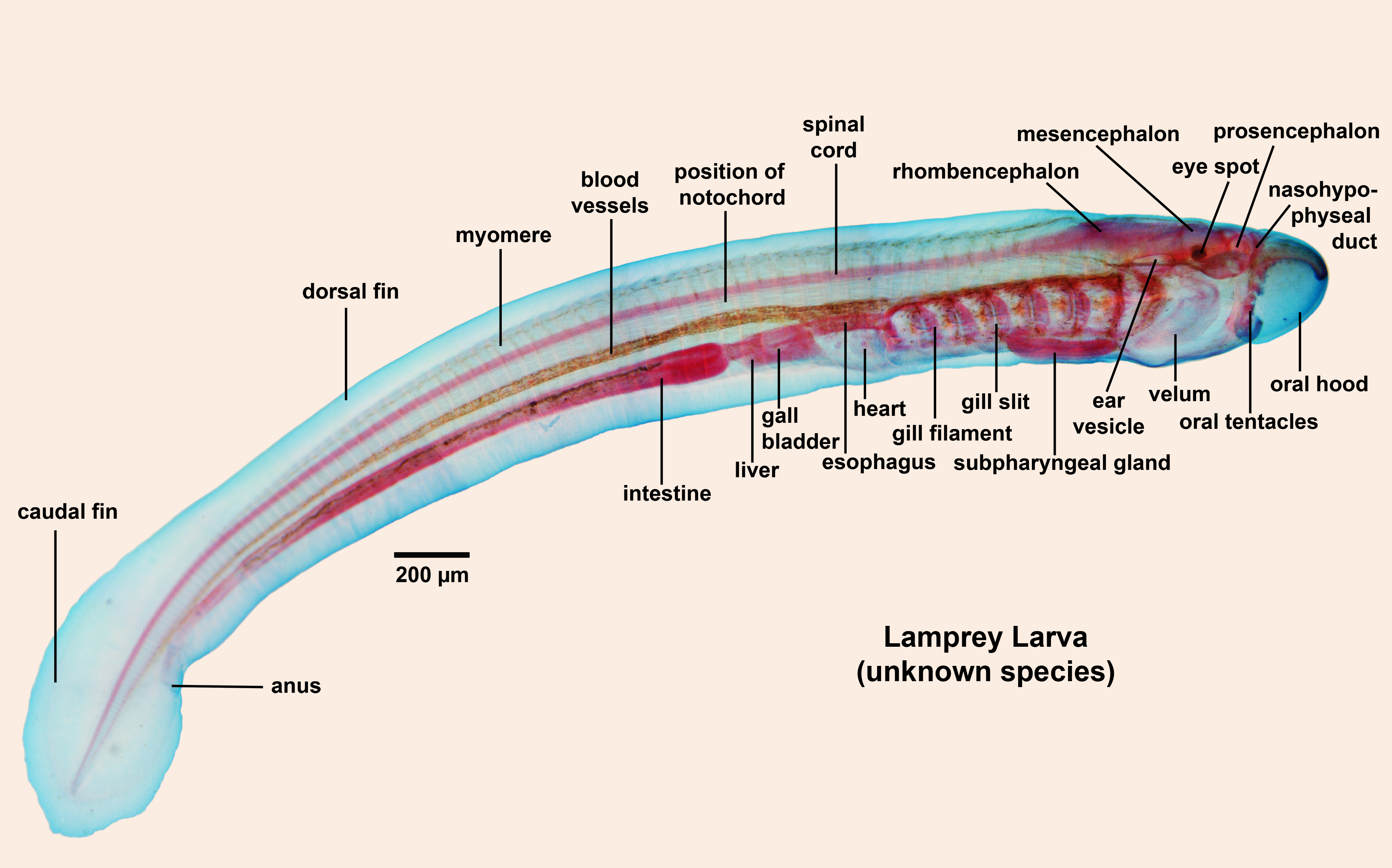
•Hyperotreti (hagfish), Hyperoartia (lampreys) and Gnathostomata are the major living groups of vertebrates.
•Their relationship is unsettled, with some favoring the "Vertebrata" hypothesis in which lampreys group with gnathostomes, and others the "Cyclostomata" hypothesis in which lampreys and hagfish are sister taxa.
•After a century of study, conodont elements were shown to belong to basal vertebrates, now named Euconodonta.
•The evolution of jawless vertebrates tracks the proliferation of bony tissue in the vertebrate body.
•Gnathostomata has a long list of derived characters, including jaws, that form from mandibular and hyoid arches. They, too, display a Hox gene duplication giving them four hot clusters.
•Placodermi has been shown to be a paraphyletic group of basal gnathostomes within which Eugnathostomata, the crown group is nested.
•For more, take GEOL431 in spring 2021.
"Say, what a lot of fish there are!"
Theodore Seuss Geisel



The Burgess Shale fossil Pikaia graciliens, known from hundreds of specimens, is often cited in popular literature as the "earliest chordate." In fact:
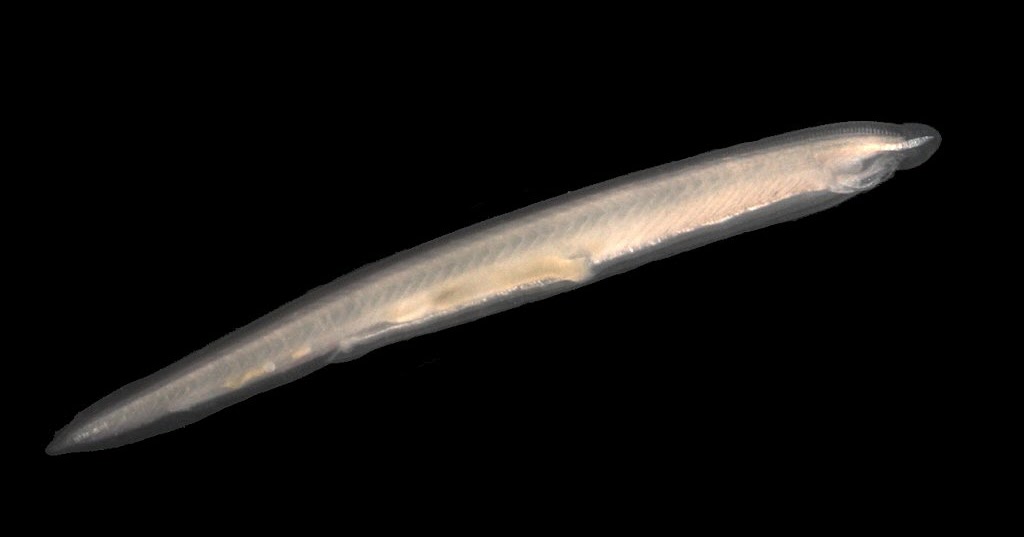
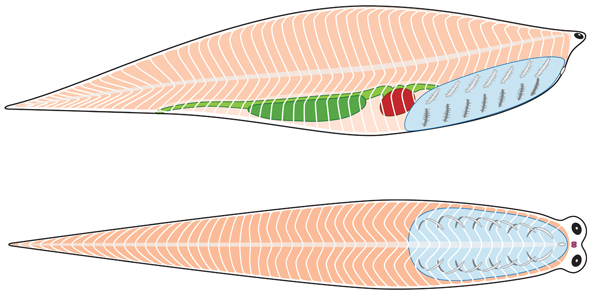
Cephalochordata: (Cam. - Rec.) "Headless fish."
Represented by the living Branchiostoma.
Confusing nomenclature: Earlier names have priority in scientific nomenclature. Unfortunately, the publication in which this animal was originally named (Branchiostoma) was largely ignored, and it was named a second time (Amphioxus). Additionally, there had been a common name (Lancelet). When the dust settled, Branchiostoma was acknowledged the proper scientific name, and amphioxus and lancelet are both considered common names. Live with it.

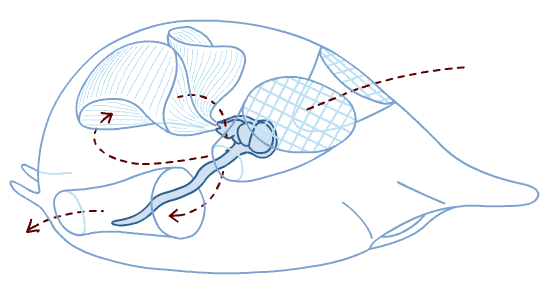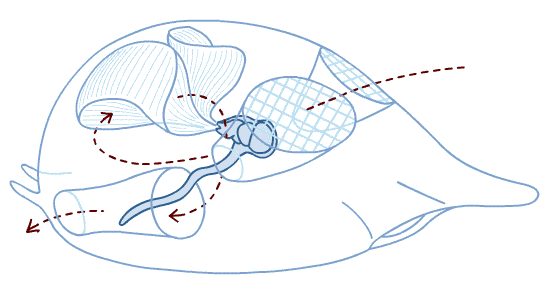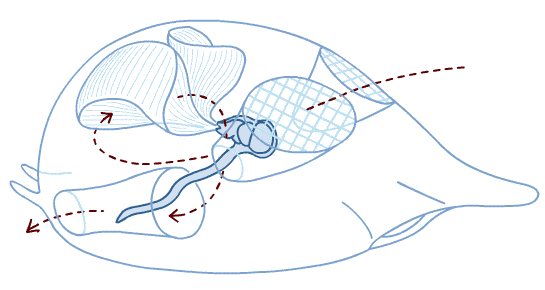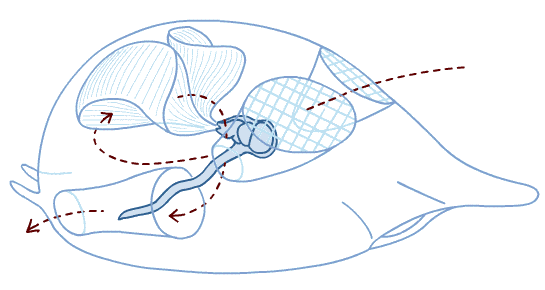
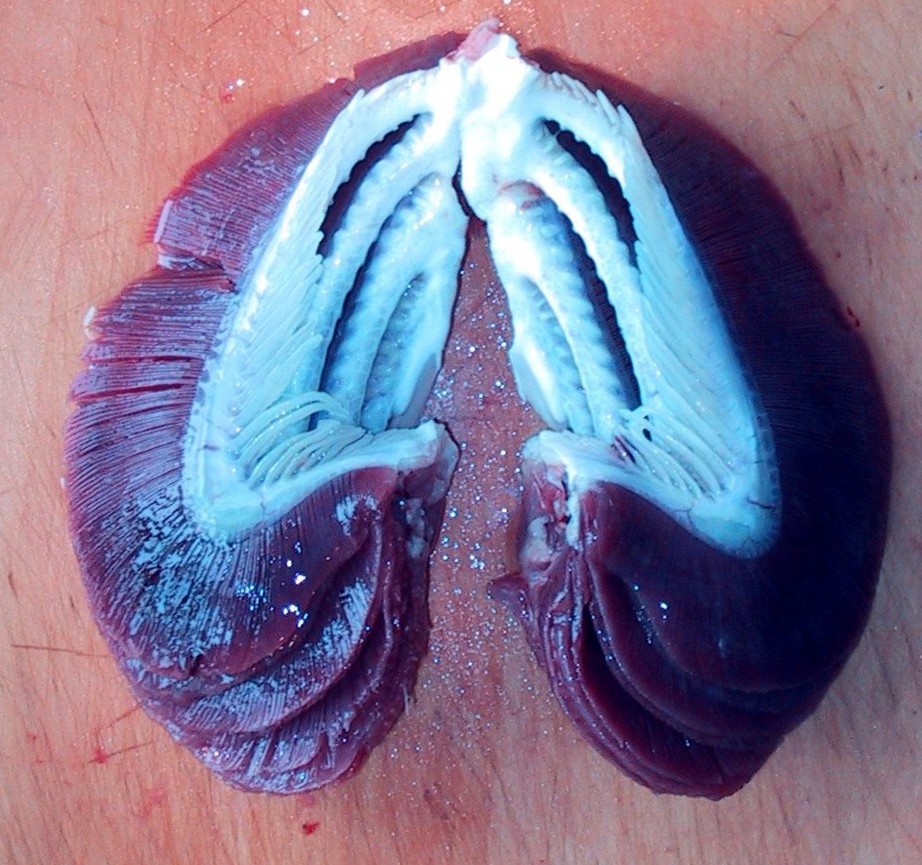
Branchiostoma ecology: Branchiostoma opens a window onto the ecology of the "swimming pharynges" at the base of Deuterostomia. It does not spend much time actually swimming, but, instead, buries itself in soft sediment, with its front end projecting into the water suspension-feeding. When disturbed, it swims in a fish-like manner, but soon settles down, digs in, and resumes suspension-feeding.
Anatomical details:
Synapomorphy of Cephalochordata:
Cephalochordate fossil record: Sparse, but present as early as the Chengjiang lagerstätte in the form of Cathaymyrus diadexus.
Interpreting cephalochordates: For decades, cephalochordates have been presumed to be closely related to Chordata. They are so close morphologically. The Wizard need only give them hearts and brains. The next group, in contrast, seems more distant.
Urochordata: (Sparse record Cam. - Rec.)
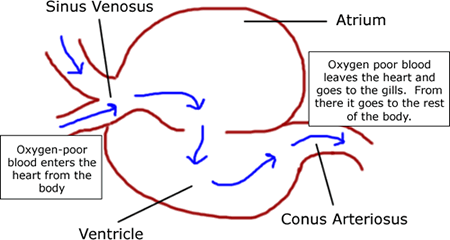
Also known as "tunicates" or "sea squirts." As adults, the body is dominated by a huge pharynx used for suspension-feeding. Adults of readily accessible species are typically attached to some hard surface, however many are free-swimming. They may be solitary or colonial.
Anatomy:
Indeed, one sub-group - Larvacea maintain their tadpole-like morphology as they metamorphose into feeding adults. (Freaky stuff - Adults secrete a gelatinous structure, the house (that's the technical term) which they inhabit and push around as they swim.
Urochordate fossil record:

When your instructors completed their PhDs, the consensus of deuterostome phylogenies held that:
Discoveries that have changed that view:
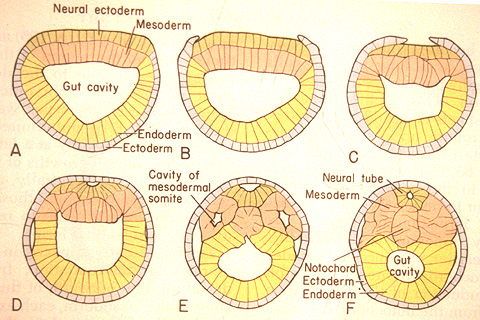
The special sense organs arise from the interaction of the neural tube with placodes of the outer ectoderm.

First question: Who is the sister taxon of "chordates with heads?"
Olfactores - Urochordate - craniate synapomorphies:
Major groups:
Craniate diversity:
First, consider the living groups:
Other characteristics:
But hagfish lack many features we see in other craniates:

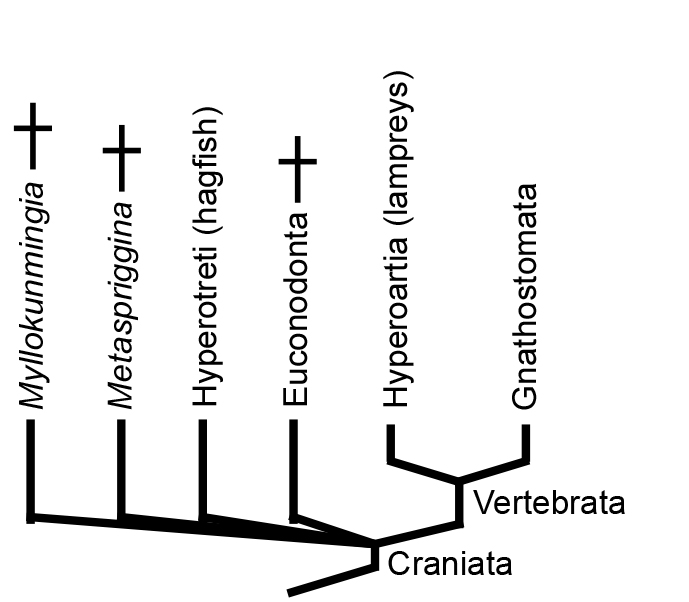
Until now we have been coy about the name of the clade containing chordates with heads. Now know the reason: Illuminating the relationship between Hyperotreti, Hyperoartia, and Gnathostomata is difficult and contentious. Two major hypotheses predominate that take their names from the positions they imply for lampreys:
This is a huge problem in vertebrate systematics that cries out for resolution. Janvier's quote is apt, but opinion is swinging toward the Cyclostomata hyopthesis. What we really need is for a Cambrian konzervat-lagerstätte to cough up a fossil basal hagfish or lamprey. Of course, maybe they have and we haven't recognized it. ;-)
In GEOL331, we reluctantly adopt the terminology of the cyclostome hypothesis.



But where does Euconodonta go on the chordate cladogram? The presence of phosphatic hard parts arguably places it, closer to Gnathostomata than to hagfish or lampreys, but there are concerns:
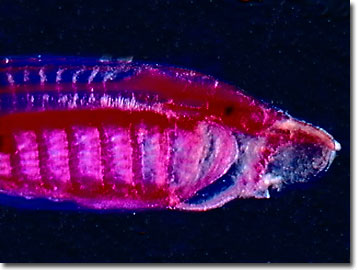
Myllokunmingia and Haikouichthys: Chengjiang gives us the best picture picture of what the ancestral vertebrate might have looked like with Haikouichthys ercaicunensis (Shu et al., 1999) H. ercaicunensis may be junior synonym of Myllokunmingia fengjiaoa). These seem to preserve:
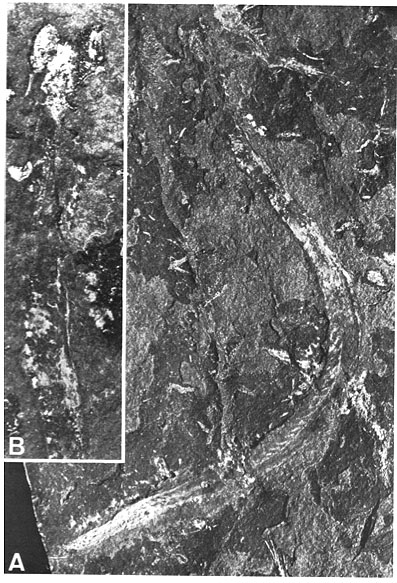
Metaspriggina: And from the Burgess shale, Conway-Morris and Caron, 2014 describe Metaspriggina walcotti (right). In this case, the presence of eucondont-like eyes and W-shaped myomeres is unambiguous.

We have such a copious record of heavily armored Early Paleozoic forms that it is tempting to forget that the group's most basal members (like Haikouichthys and Myllokunmingia) essentially lacked hard tissues. Indeed, the early evolution of Vertebrata is marked by the diversification of bony tissues and their proliferation through the body. This pattern was recently illuminated by Sansom et al., 2010 and Miyashita et al., 2019. (Composite synopsis at right.) So, we start with a review of bony evolution.

Hagfish and lampreys, as the only living jawless vertebrates, provide an interesting glimpse of early vertebrate evolution, however they lack the proper hard tissues by which we know the vast diversity of early vertebrates - bone.
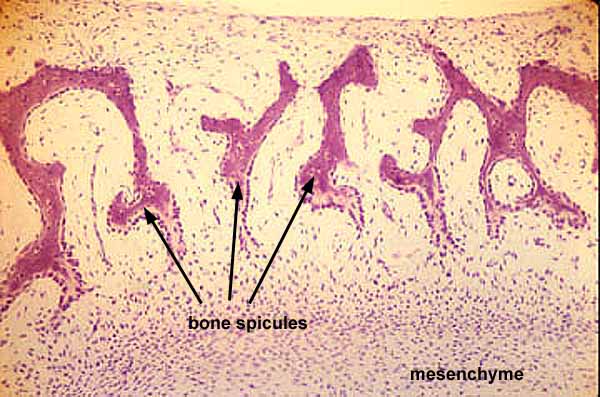
Fossil vertbrates are mostly known from hard tissues - bone and teeth. Bone is composed of:
Bone in any form only occurs among members of Vertebrata, although we know there are some vertebrates who lack it. What does the study of fossil organisms tell us about the distribution of bony tissue?

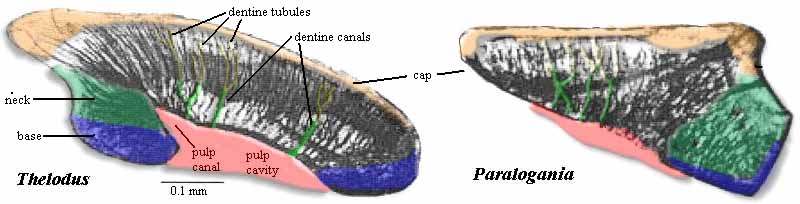
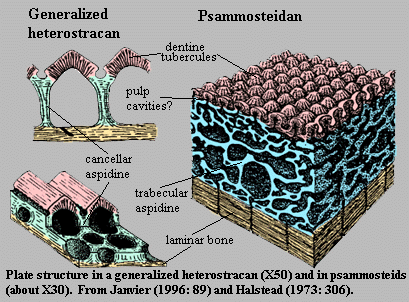
The earliest vertebrate hard tissues are small acellular elements: conodont elements, which show outer layers of enamel covering layers of dentin. Conodonts were not the only representation of craniate hard tissues in the Cambrian, however. Enigmatic, scale-like plates of bony armor called Anatolepis were also present. In this and similar creatures, histologically tooth-like denticles complete with enamel and dentin formed a composite superficial body armor.
Indeed, in many early vertebrates, there seems to have been little difference between teeth and scales, which took the form of little denticles with a pulp cavity, dentin, and enamel.
The most basal vertebrates, however, lacked any hard tissues (except for conodont elements.) A survey of early vertebrate evolution should focus on their acquisition:

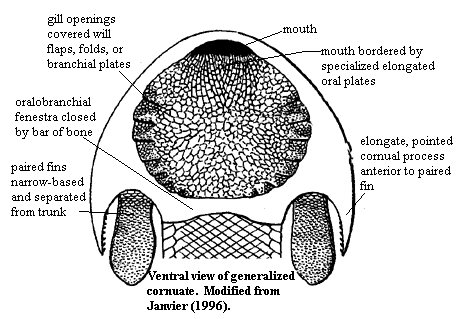
These form a clade in analysis of Miyashita et al., 2019. Illuminated by well-preserved Jamoytius and Euphanerops. These were cylindrical, vaguely lamprey-like creatures with varying amounts of acellular dermal bone plates, but without specialize mouthparts. They lack obvious adaptations to suspension feeding or to taking large prey. Deposit feeders? Many morphological details reinforce the proto-lamprey impression
Thelodonti (Ordovician - Devonian)
Morphology: Hard tissue Entirely consists of small scales that usually disarticulate when the animal dies. These scales are distinctive, consisting of enamel and dentin layers around a pulp cavity, like a vertebrate tooth. Note: from this point on the tree onward, aquatic vertebrates generally retain scales of this sort or their derivatives, regardless of any other kind of skeletal ossification they may have.
Synapomorphies with jawed-vertebrates:
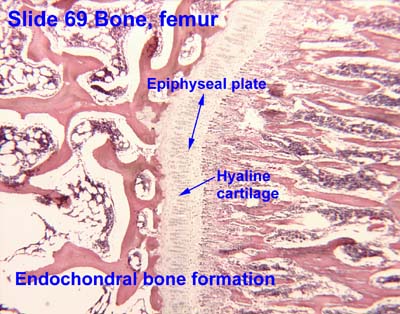
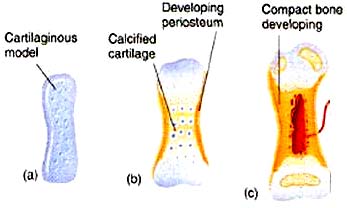
Note: Among living vertebrates, both types of ossification occur in cartilage bone, so neontologists and human anatomists use "cartilage bone" and "endochondral bone" interchangeably. Paleontologists must distinguish them, however, because perichondral ossification evolved first and many fossil vertebrates have perichondrally ossified bone without the endochondral component.

The earliest well-preserved vertebrate, the Ordovician form Sacabambaspis, ironically represents a more derived form of hard tissue, in which individual denticles are integrated into broad head-shield composite elements and joined to one another through dermal layers of aspidin, a composite of thelodont-like denticles, lamina of dentin, and cellular dermal bone. Note: It was not an internal skeleton.
Synapomorphy with jawed-vertebrates:
Galeaspida: Restricted to southern China and Indochina, then a separate continent. (Silurian - Devonian)
Morphology:
Synapomorphy with jawed-vertebrates: Perichondral bone in braincase.
Osteostraci (Silurian - Devonian): Resemble galeaspids but with differences:
Morphology:
Synapomorphies with jawed-vertebrates:
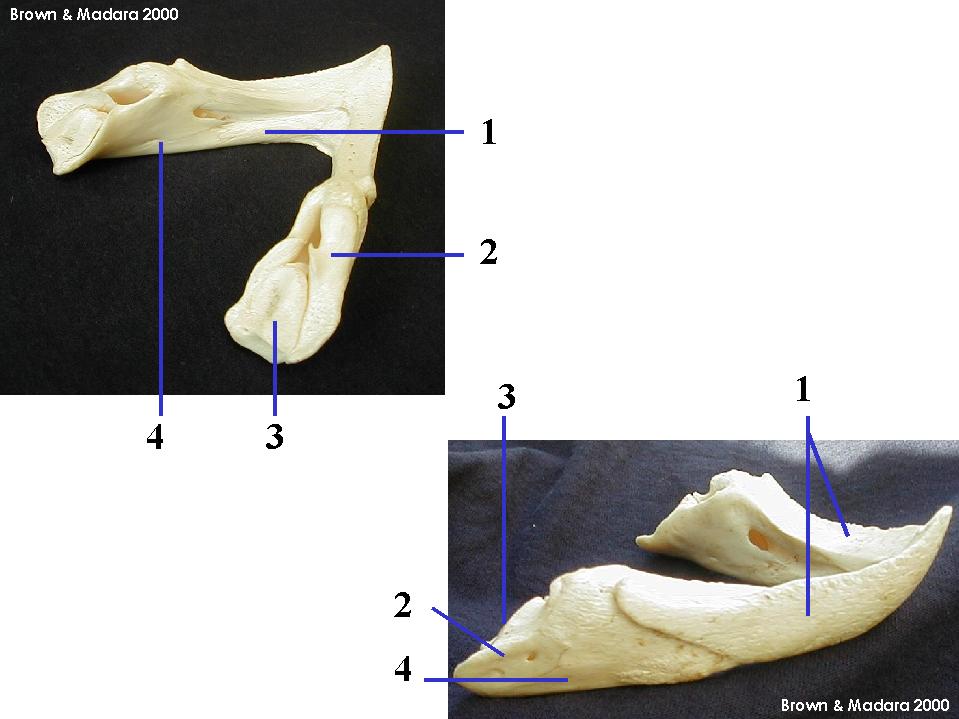
Gnathostomata: (Sil - Rec.) The jawed vertebrates. Quantum leap forward.

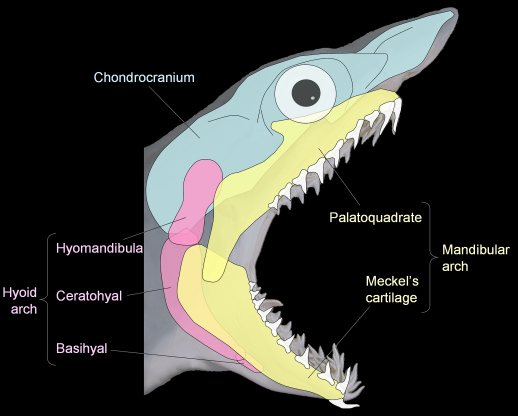
Jaws
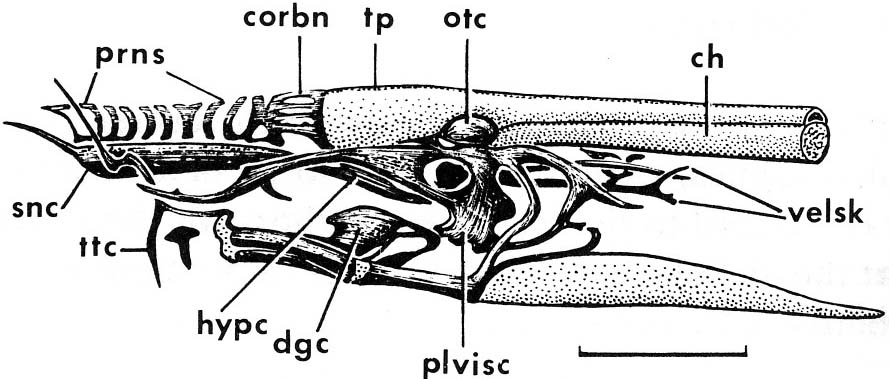
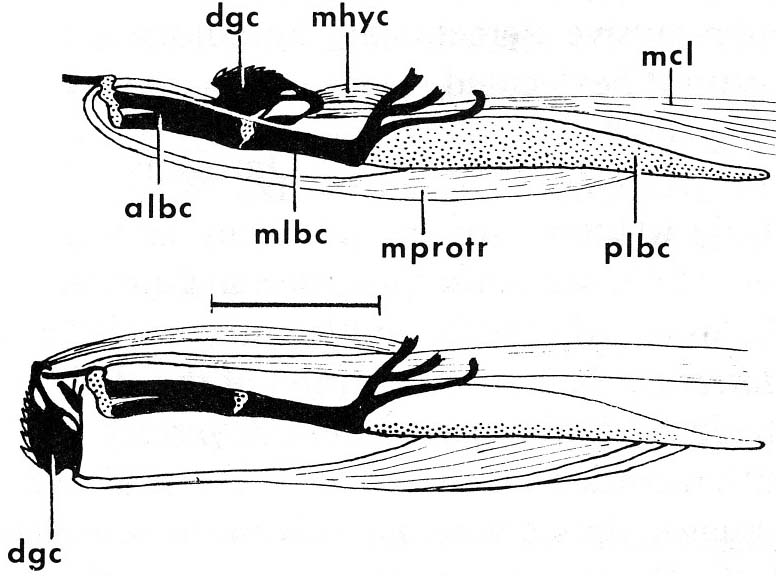
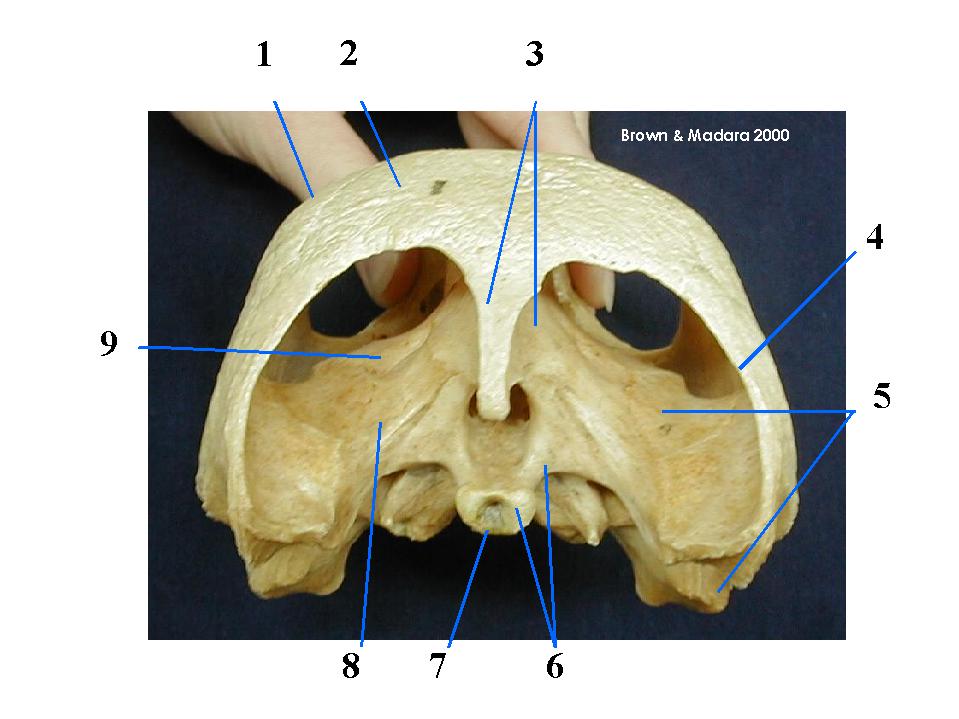
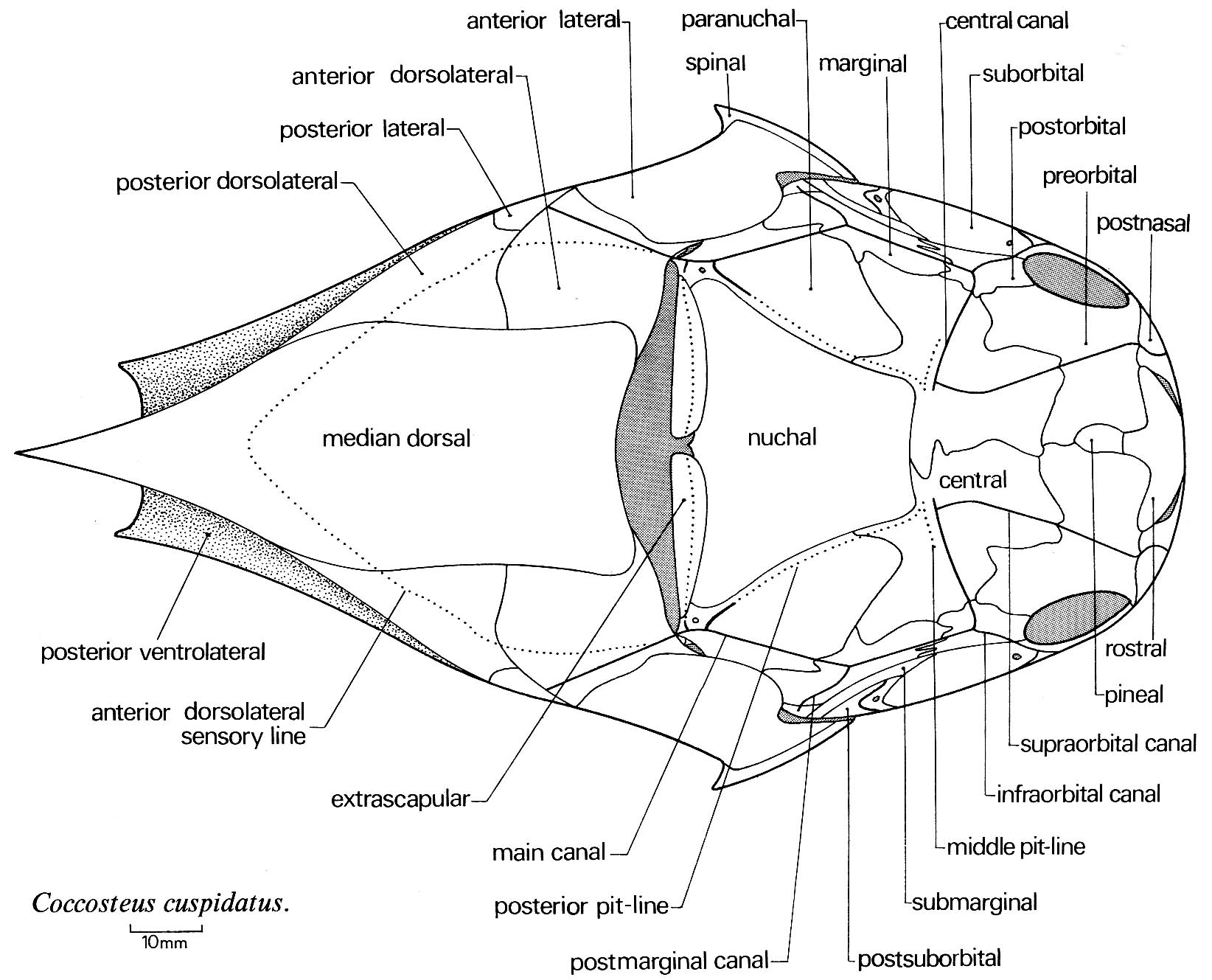
Jaw anatomy: Up until now, when we have spoken of a "skull" we have meant only a plating of dermal bone around the head, or, in the case of Galeaspids and Osteostraci, that plus in endochondrally ossified neurocranium or braincase. With the appearance of the gnathostome jaws and branchial skeleton, the skull becomes a complex composite structure.
Note: In vertebrates with bony skeletons, these components can ossify in different patterns and into different numbers of elements, but the underlying identity of the cartilage precursors remains the same.
The following illustrations show its components using the fossil bony fish Eusthenopteron as an example.




Note, whereas the branchial arches are separated from the hyoid arch and from one another by gill slits, the space between the hyoid and mandibular arches is either closed or perforated by the spiracle. (This is particularly conspicuous in rays, who breath in through it.)

Traditionally, three major groups of unknown relationships were recognized:
Placodermi: Silurian to Devonian armored gnathostomes experienced a rapid worldwide diversification and sudden decline.


Eugnathostomata: The gnathostome crown-group. The last common ancestor of Chondrichthyes and osteichthyes and all of its descendants.
Synapomorphies of Eugnathostomata include:
All organisms more closely related to modern sharks and chimaeras than to any other gnathostomes.
Synapomorphy: Prismatic calcification of the cartilage. Chondrichthyans do not lack other hard tissues. Various groups make teeth, fin spines, and dermal armor out of dentine and enamel.

Synapomorphies:

This creature has features seen in both primitive sarcopterygians and actinopterygians, but retains others typical of primitive stem osteichthyans and stem gnathostomes, such as the stout pectoral spine.
Actinopterygii: (Dev - Rec)
The vast majority of "fishlike" bony fish.
These have so vast a record from the Devonian on that it's hard to know where to start. Alas, they exceed the scope of the course.
Sarcopterygii: The lobe-finned fish and tetrapods.

Tetrapod in brief. A survey of Tetrapoda, including all land vertebrates and some secondarily aquatic ones, exceeds our scope. Don't despair. There are many sources of information. Major living tetrapod groups include:
To Next Lecture.
To Previous Lecture.
To Syllabus.


/larvacean-5a3ad88fbeba33003737d0a3.jpg)












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}