Fall Semester 2018 Variation and Fossil Individuals
Birds of a different color: Three major genes set feather hue in pigeons. 2014. Phys.org.
Key Points:
•The individual is the only true unit of variation among living things.
•The recognition of variation as an intrinsic part of nature was instrumental to the development of the Theory of Evolution.
•Variation results from a range of biological factors including: Individual, ontogenetic, sexual, generation-alternation, and ecophenotypic.
•Taphonomic issues impose their own sources of variation in fossils.
•Different taxa employ a range of growth strategies which impart different styles of ontogenetic variation
•Biomechanical constraints impart variation as organisms scale allometrically
•Heterochrony represents the linkage of ontogeny (growth) an evolutionary change.
•The grouping of variable individuals into species is problematic:
•Asexual "species," hybrids, hybridogens, morphoclines, sibling species, ring species, and closely related species clusters frustrate our understanding.
•Competing species criteria, including the Biological, Evolutionary, and Phylogenetic Species Concepts would divide up organismal diversity differently.
•Depending on the tempo and mode of evolution, the chronological duration of species is similarly uncertain.
"THE MANY slight differences which appear in the offspring from the same parents, or which it may be presumed have thus arisen, from being observed in the individuals of the same species inhabiting the same confined locality, may be called individual differences. No one supposes that all the individuals of the same species are cast in the same actual mould. These individual differences are of the highest importance for us, for they are often inherited, as must be familiar to every one; and they thus afford materials for natural selection to act on and accumulate, in the same manner as man accumulates in any given direction individual differences in his domesticated productions.
(Charles Darwin, 1859. The Origin of Species)
Sources of Variation within Populations:
The ultimate goal of taxonomy is the description of the diversity of living things. Without this knowledge, we can barely begin to understand the fossil record, however before attempting to do taxonomy, we must consider the mechanisms that generate diversity among organisms.
A small sample of human variation
Individual: Darwin's most revolutionary thought - the abandonment of the typology of Plato and Aristotle for the reality of individual variation. Upon reflection, the wonder is not that individuals are distinct. Rather, it is that that variation tends to cluster together in an internested hierarchy of groups.
Our modern perspective:
Individuality results from the shuffling of genes into novel combinations by the processes of sexual reproduction, accompanied by the occasional mutation.
The internested hierarchy results from patterns of common ancestry implicit in the idea of evolution.
Organisms need to function at all stages of ontogeny (individual development) but don't have to look similar look similar.
In some, development is essentially direct (E.G. Pachrhinosaurus right). In others, ontogenetic stages maybe morphologically and ecologically so different that members of the same species are difficult to recognize. (E.G. blue crab life stages)
And yet, extrapolating from the Malthusian principle of reproductive excess, we should assume that most of the individuals that we find as fossils don't represent full adults.
Sexual: Sexual morphs can be anything from indistinguishable to utterly dissimilar. One thing is certain, even with fossils. The organisms had to come together to mate, so we would expect sexual morphs to inhabit the same time and space.
Alternating generation: Popular culture introduced the alternation of generations through the Alien film with alternating face-hugger and chest-burster generations. Surprise! This happens frequently in nature, including in animals and plants. Recognizing alternating generations as members of the same species is difficult with living taxa and effectively impossible with fossils unless they die in the act of propagating and are preserved.
Acropora ecomorphs from below (left) and above (right) wave-base
Ecophenotypic:
Organisms of the same species may look substantially different depending on their environment. E.G. Scleractinian corals in different energy regimes. (right). A common issue with plants and colonial animals, but occasionally with others. The axolotl, a Mexican lake salamander, for example typically suppresses metamorphosis but will metamorphose under stress conditions.
A contrived example - The same trilobite alternately preserved.
Taphonomic/Preservational Not a source of variation that neontologists generally worry about, but paleontologists always must.
Specimens of the same species that have undergone different taphonomic processing might look quite different (right).
Chimaeras - The creation of fictional organisms that are composites of the remains of representatives of different species whose fossils came to rest in the same place are disturbingly common. A high-profile example was the Chatterjee, 1991 description of Protoavis, ostensibly a Triassic bird-like theropod but more likely a chimaera.
Growth Strategies:
After individual variation - an essential driver of evolution, ontogenetic variation is arguably the most significant source of diversity, so its interpretation merits detailed attention. Interpreting ontogenetic status requires first that the growth mode of hard parts be identified. The major growth strategies:
Gastropod shell with growth lines
Marginal accretion: (E.G. cnidarians, brachiopods, mollusks, some vertebrate elements): Growth occurs by deposition of hard tissue at the periphery of existing hard parts. This typically leaves "growth lines" that serve as a record of the element's previous size and shape.
Addition: (E.G. Foraminiferans, internal structures of cephalopods, echinoderms): Discrete new parts added or intercalated, with little change afterward. Shelled cephalopods, for example, sequentially add internal shell chambers (right).
Bryozoan zooecia
Serial Addition in colonial organisms (cnidarians, bryozoans): A special case of addition where the parts replicated are skeletons of zooids comparable to those of entire other organisms.
Ecdysis (a.k.a. Molting) (arthropods): each growth stage or instar is entirely new
hard material part.
This allows for radical transformation between growth stages. This is particularly extreme in insects (E.G. winged cicada adult emerging from skeleton of burrowing nymph, right)
leaves discrete size classes representing age cohorts.
Allows a single organisms to leave many potential body fossils.
Schematic cartoon of vertebrate bone growth
Continuous Modification (vertebrates): Vertebrates experience a continuous remodeling of skeletal tissues:
Skeletal elements are pre-formed in membrane or cartilage, that serves as the template for the deposition of bone, and is often replaced by it.
Bone, once laid down by specialized cells called osteoblasts, is remodeled throughout ontogeny by and removal by osteoclasts coupled with ongoing deposition. This typically obscures details of earlier life stages.
But note that many creatures combine strategies. E.G.:
A distinct ontogenetic issues is that of scaling. Typically, different parts of the organism grow at different rates - i.e. they experience allometric scaling. This results from a simple fact of geometry:
Surface area scales as the 2/3 power of volume when an object is scaled up isometrically - with all dimensions scaling at the same rate.
Thus, if we double the scale an object in all dimensions (i.e. we scale it isometrically), the load experienced by any part of its load-bearing surface increases by two. If we keep doing this, the material will reach its mechanical limits and break under the load.
Organisms cope with changes in the mechanical forces they experience during growth by scaling allometrically - with different parts of the body growing at different rates:
Positive allometry: a feature scales up faster than than others.
Negative allometry: a feature scales up more slowly than than others.
Consider...
A small Allometrosaurus (1) scales up isometrically (2) or allometrically (3)
To compensate for changes in mechanical loading, body proportions must change as the object scales up. In the natural world, organisms scale allometrically (i.e. in unequal proportions as they grow, in large part to maintain constant levels of loading on their tissues. In the example at right, the leg bones of a little Allometrosaurus support its body well, but when scaled up isometrically by a factor of two, its leg bones are dangerously overloaded. To compensate, the width of the leg bones must scale with positive allometry (3) so the load experienced by each parcel of bone does not change.
This form of allometric scaling is called scaling with mechanical similarity.
White-tailed deer and cape buffalo - scaling with mechanical similarity
Allometric scaling is also evident in comparisons across taxa.
Small and large members of the same group such as the white-tailed deer (left) and cape buffalo (right), both members of Artiodactyla, may have different limb proportions but scale with mechanical similarity in order to move about in basically similar ways.
Of course, this has limits, too. If the critter's legs scale up faster than its heart and lungs, it will eventually be unable to support them. Thus, large animals typically behave differently than small ones to limit loads and conserve energy. An elephant moving full-speed is still walking, whereas a small mammal can afford to bound with all four feet in the air.
The "evolution" of Babylon 5's Jason Ironheart
Growth and evolution:
Remember, ontogeny pertains to the individual, evolution to the lineage. (TV sci-fi plots notwithstanding.) That doesn't mean they aren't connected in any way. Now that we know something about how organisms grow, let's consider how alterations in ontogeny over evolutionary time can drive evolutionary change through the process of heterochrony.
Heterchrony:
Subtle changes over evolutionary time in an organism's developmental timetable are a potent source of overall evolutionary change. This is an idea with a history as long as the study of evolution.
Ernst Haeckel (1836 - 1919) Noted that the embryos of evolutionarily derived creatures may pass through stages in which they display ancestral traits. The human embryo, for instance, at certain stages has a tail and open gill slits. Proposed what he termed the biogenic law which maintains that "ontogeny recapitulates phylogeny." He attempted to integrate it with Darwinian evolution through the notion that evolutionary change is accomplished by the addition of new changes to the end of the embryological or developmental sequence. This idea seems simplistic when you consider what it implies about the evolution of butterflies, but it was eagerly embraced for the first half of the 20th century. (Note: Haeckel was an interesting originator of vocabulary. He gave us the terms "heterochrony" and "biology."
Karl Ernst von Baer (1792 - 1876) - Was an opponent of recapitulationism and set forth a series of developmental laws that held that:
Development is a process of differentiation from generalized to specialized form, with general structures appearing first.
The embryo of any creature is never identical to the adult of any other.
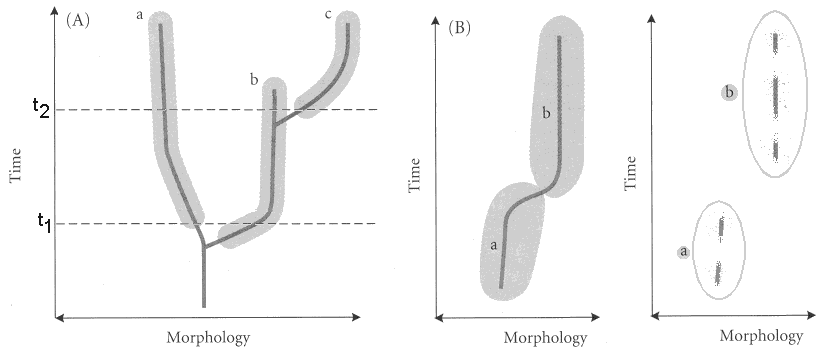
Stephen J. Gould: (1941 - 2002) Accumulating evidence during the 20th century revealed many weaknesses in the "biogenic law." Von Baer's laws, while holding up intact, cried out for elaboration. In 1977, Stephen Gould stepped to the plate in the book Ontogeny and Phylogeny with the terminology and organization of heterchrony as we understand it. We have two basic forms of heterochrony, and three distinct parameters that can be tweeked to produce the change. The result is six distinct varieties. Study the diagram below for full illumination.
General patterns of heterochrony:
Adult derived axolotl (left) resembles juvenile ancestral tiger salamander (right) from The Planted Tank and Axolotls
Paedomorphosis: The adult of a derived organism resembles a juvenile of the ancestor. Compare the derived, paedomorphic axolotl to a "normal" relative, the tiger salamander. (Adult tiger salamander.)
Juvenile derived veiled chameleon (left) resembles adult ancestral common chameleon (right) from FLchams and Wikipedia
Peramorphosis: The juvenile of a derived organism resembles an adult of the ancestor. Compare a the derived peramorphic veiled chameleon
to a "normal" ancestral common chameleon.
Gould added clarity to these terms by identifying the developmental parameters that could be tweaked to produce either major pattern, and naming the different pathways that resulted. In Gould's system, heterochronic change could result from changes in:
Rate of development.
Time of onset of development.
Time of offset of development.
Combining these developmental parameters gives us:
Varieties of Paedomorphosis:
If this is the ancestral life history:
Neoteny: Reduced rate of development
Postdisplacement: Postponed onset of development
Progenesis: Developmental offset occurs at an earlier time.
Varieties of Peramorphosis:
Acceleration: Increased rate of development
Predisplacement: Onset of development occurs at an earlier time
Hypermorphosis: Delayed developmental offset.
Cultural "evolution": Note that paedomorphosis and peramorphosis occur in popular cultural figures.
Quite frequently, a logo or cartoon character will be juvenilized so that it will be "cute." E.G:
All of this has huge consequences for Morphometrics, the quantified statistical comparisions of shape.
But underlying all of this is a difficult philosophical issue:
Fossil Species: Species Concepts and Criteria
Species (pl. "species"; Latin for "kind"): generally considered the fundamental
unit of biological diversity. Certainly is the primary entry in databases of diversity,
abundance, occurence, etc. from modern and fossil assemblages, ecosystems, etc.
But, WHAT ARE SPECIES?!? We (sometimes) know them when we see them, but how do we
recognize them? What is our species criterion?
This is known in biology as the "species problem".
Difficulties with species delineation:
"Species" are our attempt scientifically to codify traditional "kinds," populations of interbreeding critters that are more or less morphologically uniform. Seems easy, but when you scrutinize living diversity in detail, a number of problems come up:
Asexual "species:" Many creatures reproduce primarily or entirely asexually. How do species concepts apply to them?
Hybrid Galápagos iguana
Hybrids: Creatures belonging to morphologically and behaviorally distinct groups can become "confused" and mate with the wrong partner, especially when normal ecological or geographic barriers break down. (E.G. the hybrid of the Galápagos Land and Marine iguana shown at right.) In nature, this is far from unusual, and they span a broad spectrum from clearly maladaptive sterile hybrids to fertile ones.
To see how weird things can get, consider the case of hybridogens such as the molly Poeciliopsis viriosa from Sonora. In this morphologically distinct "species," each individual is female and gets her paternal genome through her mother's mating with males of at least two closely related normal species.
Thus, hybridization spans the range from extremely maladaptive to essential to the perpetuation of hybridogenic "species."
Sibling species: Sister taxa (i.e. descendents form a recent common ancestor) that are morphologically indistinguishable, but distinguished by molecular, karyological, or behavioral traits. E.G. stonebashers of the genus Pollimyrus, who tell one another apart by difference in their calls.
Morphoclines: When interbreeding populations occupy a large range, they often display visible morphological gradations. For example, the Northern Flicker (Colaptes auratus) occupies most of North America and grades from the "yellow-shafted" form in the East to a "red-shafted" form in the west.
Ring species: Species spanning a broad morphological continuum across a wide geographic range in which members of a local population can interbreed with members of adjacent populations, but end members of the continuum cannot interbreed.
Offshore (left), transient (center), and resident (right) orcas.
It gets worse: Even without these complications, the delineation of "species" can be daunting. Consider the killer-whale Orcinus orca: Three morphological types are known from the Pacific Northwest (above) that are behaviorally distinct. and not observed to interbreed.
"Residents" are sedentary fish-eaters.
"Transients" are nomadic marine-mammal hunters.
"Offshores" are nomadic fish-eaters.
Nine such populations are known worldwide. Is Orcinus orca a species with distinct sub-populations, or a cluster of closely related species?
Species criteria:
The paradox of species criteria: We humans feel compelled to define species precisely when, in nature, their boundaries are fuzzy, indistinct, and best described probabilistically.
And yet, living things do seem to group into morphologically distinct populations rather than grading across wide ranges of morphospace without breaks, even those that reproduce asexually. Traditionally, species are morphospecies: "a diagnosible cluster of individuals within
which there is a pattern of ancestry and descent, and beyond which there is not." However,
individual variation is a basic attribute of ALL organisms; and geographic variations are
very common as well. At what point are two different geographic populations different at "the
species level"? Your instructors' personal rules of thumb:
Merck: Two populations represent distinct species when they are sufficiently different that hybridization between their individuals reduces the fitness of their offspring.
Holtz: Any species concept that requires us to view lions and tigers as belonging to the same species is a bad one.
But we're not the experts. Let's refer to the view of Charles Darwin, who felt that species were merely well delineated varieties whose distinctiveness from other organisms arose from the fact that intermediate forms were now extinct.
None of this has prevented biologists from attempting to develop hard and fast species concepts. Major attempts at species definitions that have gained significant traction include:
Kekaimalu the wholphin
Ernst Mayr's Biological Species Concept (BSC) (1942): "a species is an array of populations
which are actually or potentially interbreeding, and which are reproductively isolated from
other such arrays under natural conditions." Problems:
Reproductive isolation is meaningless in asexual organisms
has not been tested for the vast majority of living organisms
fails to account for the truly vast numbers of morphologically distinct populations that are prevented from interbreeding simply by geography or ecology, and may do so under altered circumstances. (E.G. Wholphin (right), improbable offspring of captive bottlenose dolphin and false killer whale.)
is impossible to test for fossils (including fossils which we would otherwise include within modern species)
ignores that fact that reproductive isolating mechanisms (distinctive calls and displays, mechanical or karyological barriers to successful mating) are evolved responses to the challenge of reduced fitness in hybrids, and may not be present in distantly related species for whom hybridization is unlikely to occur anyway.
But the Biological Species Concept possesses one great strength: It at least suggests a falsification criterion for the hypothesis that two populations are distinct species: Can they interbreed?
George G. Simpson's Evolutionary Species Concept (ESC) (1961): "An evolutionary species is a lineage evolving separately from others and with its own unitary evolutionary role and tendencies." Philosophically free from the difficulties of the Biological Species Concept, but lacking clear criteria for testing actual hypotheses of species identity.
Niles Eldredge and Joel Cracraft (1980) Reformulated the ESC
after the cladistics revolution into the Phylogenetic species concept (PSC): Regarded a species as the smallest diagnosable taxon having an origin in a lineage splitting event. In this scheme, "a diagnosable cluster of individuals within which there is a parental pattern of ancestry and descent, beyond which there is not, and which exhibits a pattern of phylogenetic ancestry and descent among units of like kind." This offers the philosophical rigor of the ESC with the practical recognition criterion of diagnosability - it is possible to recognize through a combination of derived character states.
Warning! Don't think that these criteria are simply different ways to reach the same underlying truth. In many cases, their practical application might be the same, but they are different concepts. Consider pantherine cats: In the wild, lions, leopards, jaguars, etc. are morphologically and behaviorally distinct, but in captivity they produce fertile hybrids frequently.
By the BSC, Panthera is a species with highly variable morphotypes.
By the PSC, Panthera is a higher-order group of closely related species.
Paleontological applications: There have been many minor variations on these themes. For paleontologists, however, the Phylogenetic Species Concept is most frequently employed, simply because we just can't test reproductive isolation in fossil taxa. We can, however, test hypotheses of phylogeny - the branching pattern of evolution, in such a way as to determine whether individuals might have belonged to single lineages.
Caveats:
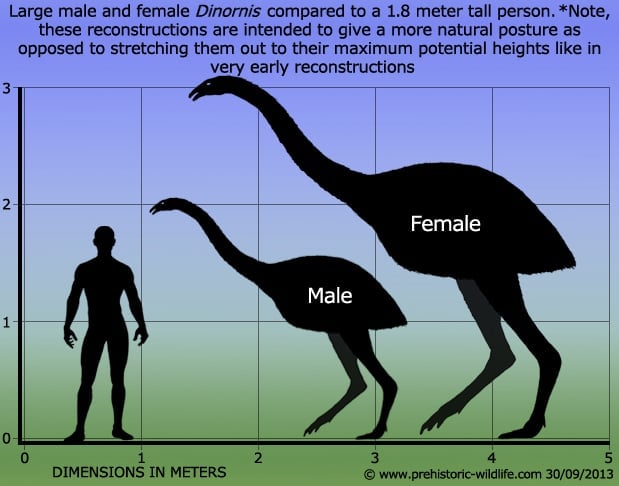
Some degree of similarity is necessary in fossils before the thought enters our heads that they might belong to the same species. E.G.: Dinornis robustus a particularly large extinct Moa, and Dinornis struthioides a small and slender one were considered distinct species (albeit sympatric) until genetic analysis revealed that all D. robustus were female and all D. struthioides were male. Oops.
One case where paleontologists might approximate a test of reproductive isolation focuses on the display structures that the critters, themselves, used to know with whom they should be mating. The Specific Mate Recognition Concept bases species identification on these features. Not applicable in many cases, but foolish to ignore when it's there. Examples: Coke's hartebeast and sympatric topi. Cranial display structures in ceratopsid dinosaurs.
Speciation: The Origin of Species
So far, we have only considered the present time slice. When we look at the past, other issues rise up. Remember that despite the foregoing, "paleospecies" are necessarily morphospecies. Whatever species concept we favor, we can't absolutely assess their mating habits.
Just as species boundaries are fuzzy in space, they are fuzzy in time. When does a species begin and end? In some cases, we can definitely say:
Speciation events in which one evolving lineage splits into two mark the beginning of the two daughter species.
Extinction events mark the end of a species.
But we must account for more than that.
Two major models of species origins in geologic time:
Anagenesis: fossil populations changing from one form to another gradually
Suggests sympatric speciation
"Extinctions" of any member successional "species" would be pseudoextinction: an
arbitrary division of the anagenetic series; "originations" would be similarly arbitrary
And yet, evolving lineages may undergo pulses of evolution that make the identification of segments of the lineage possible. These are called chronospecies and are, by necessity, somewhat arbitrary.
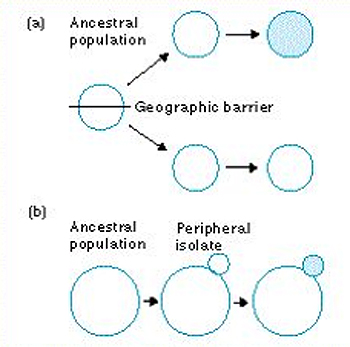
Cladogenesis: Origination of species through speciation - i.e. lineage splitting event.
Consistent with allopatric speciation as the result of the appearance of a geographic barrier to gene flow or the peripheral isolation of a population. (E.G.: The Hawaiian goose is a peripheral isolate of the Canada goose.)
Gives discrete origination point (the cladogenetic event)





























 All of this has huge consequences for Morphometrics, the quantified statistical comparisions of shape.
All of this has huge consequences for Morphometrics, the quantified statistical comparisions of shape.





 Anagenesis: fossil populations changing from one form to another gradually
Anagenesis: fossil populations changing from one form to another gradually















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}