Fall Semester 2020 Animals with Lophophores: Bryozoans & Brachiopods
Terebratulid brachiopods - Not dead yet!
Key Points:
•Traditional Lophophorata includes bryozoans, brachiopods, and phoronids - animals with lophophores. Monophyly uncertain. Some animals with lophophores - Pterbranchia - have never been considered closely related.
•Phoronids are closely related to brachiopods, but lack valves.
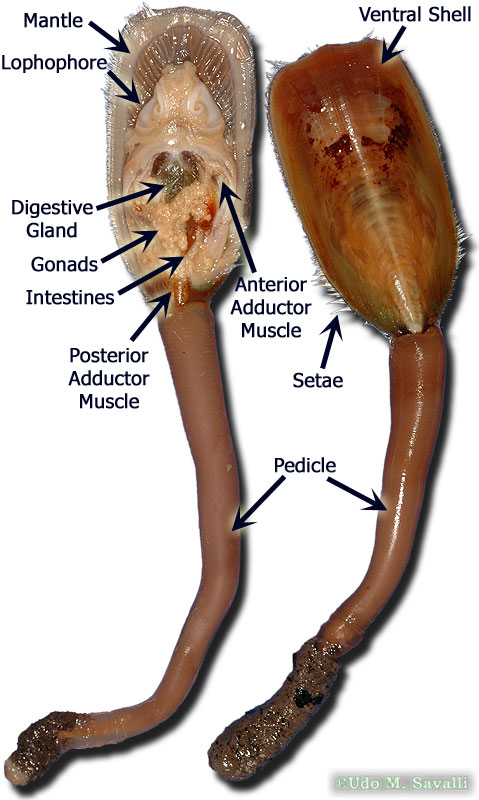
•Brachiopods have a characteristic morphology with peduncle, lophophore, and two valves. Most are sessile epifaunal suspension-feeders.
•Brachiopod have an extensive traditional taxonomy. Phylogenetic analysis is beginning to make inroads, revealing three major groups: Lnguliformea, Craniiformea, and Rhynchonelliformea.
•The traditional "articulate," brachiopods belong to Rhynchonelliformea. Sometraditional taxa are monophyletic, others are not.
•Brachiopod evolutionary trends show first appearance in Early Paleozoic, peak diversity during the Late Paleozoic, and reduction to marginal environments in the Mesozoic and Cenozoic.
•Creatures on the brachiopod stem include the soft-bodied Drakozoon, Micrina from the Tommotian small-shelly fauna, and, apparently, hyoliths.
•The oldest crown-group brachiopods, including Xianshanella haikouensis and Askepasma saproconcha are from the early Cambrian.
•Bryozoans are colonial and display characteristic zoarium (colony) and zooid morphology/anatomy.
•Bryozoans can't eliminate nitrogenous waste, instead storing it in their tissues. They then can "eliminate" their internal tissue and regenerate from their body walls.
•Bryozoans include groups with and without substantial fossil records, each with characteristic diagnostic features.
•The Ordovician, Siluro-Devonian, Late Paleozoic, Late Mesozoic, and Cenozoic each display a characteristic record of distinct bryozoan groups' apex zones. Each major extinction event pruned them significantly, with recovery from the end-Permian event being especially prolonged.
•Lophophorate systematics are currently unsettled, however recent work points toward a monophyletic Lophophorata.
"A popular belief has developed that a lifetime of research is the essential qualification for identification of bryozoans."
(W. H. Twenhofel and R. R. Shrock, 1935.)
As discussed previously, the monophyly of Lophotrochozoa is well supported by molecular and morphological evidence. We now consider "Lophophorata" animals with ciliated feeding appendages called lophophores. Note: there are other animals with lophophore-like appendages that we aren't considering, including:
Bryozoa: (AKA "Ectoprocta." Ord - Rec.) Tiny colonial "moss animals." Secrete colonial skeletons (either organic or calcareous). Distinguished by their anuses being outside the base of their hollow lophophores.
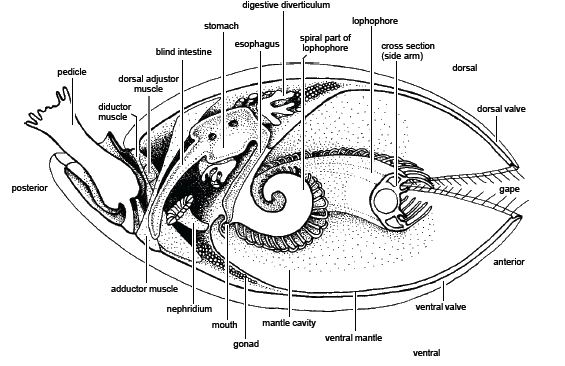
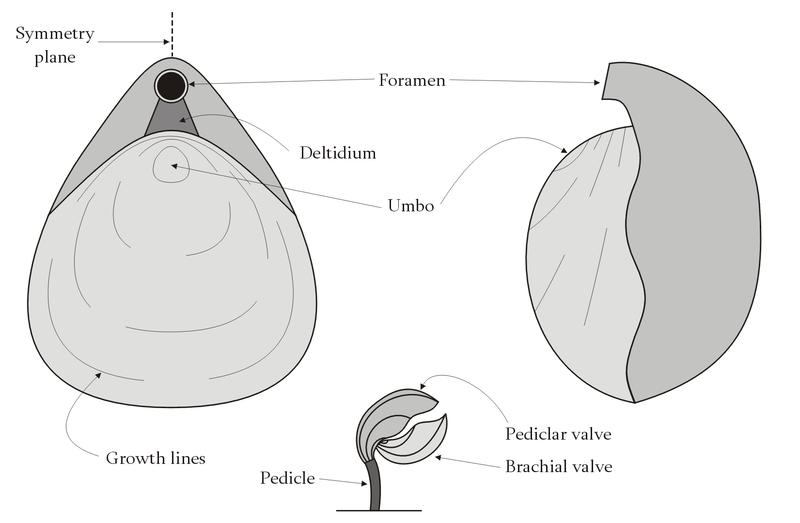
Brachiopoda: (C - Rec.) "Lamp-shells." Macroscopic solitary bilaterians that attach to the substrate with a pedicle (stalk) and are protected by two calcareous or chitinophosphatic valves (shells).
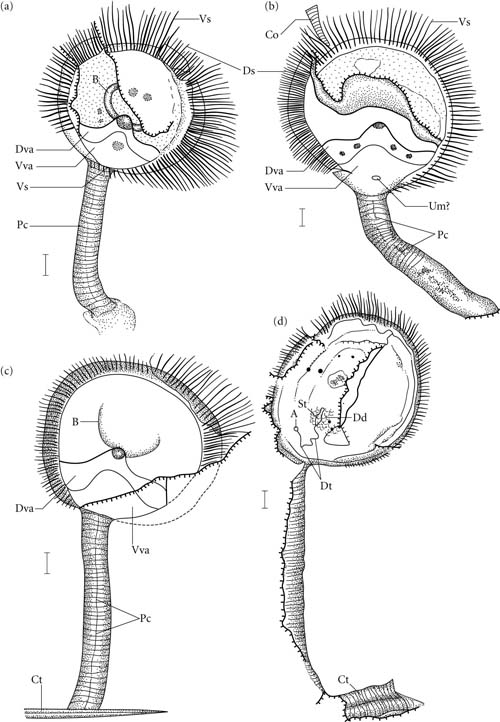
Phoronida: No unambiguous fossil record. Soft, burrowing, worm-like creatures that project their unprotected lophophores into the water column to feed.
Entoprocta: (C - Rec.) Solitary or colonial zooids superficially resemble bryozoans, but with solid (i.e. non-hollow) lophophores that encircle both mouth and anus. There is no coelom, and colony skeleton is chitinous. Unambiguous fossil entoprocts date from the Jurassic, but Zhang et al. 2013 have identified the solitary Chengjiang taxon Cotyledion tylodes as an entoproct. Opinion is divided about the Burgess Shale taxon Dinomischus.
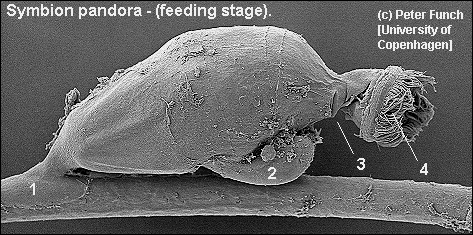
Symbion pandora
Cycliophora: (No record) Ectoparasites living on the mouthparts of lobsters, first described by Funch and Kristensen, 1995. Virtually unknown.
The relationships of these groups are discussed below. Mostly, we are concerned with Bryozoa and Brachiopoda, which have substantial fossil. records.
Phoronid anatomy
Phoronida:
(No record) Represent what we might imagine to be the ideal ancestral lophophorate. Characteristics:
Always solitary
Macroscopic
Infaunal benthic marine
Possesses
Nephridia for waste excretion
Closed circulatory system with blood cells and hemoglobin (no distinct heart)
Why do we,as paleontologists, care about critters with no fossil record? Because they illuminate the relationships of critters with good records. Consider Conway-Morris and Peel, 1995's notion of a sister-taxon relationship between brachiopods and the early Cambrian (Sirius Passet) taxon Halkieria. Sounded reasonable only as long as:
you ignored phoronids
you weren't aware of synapomorphies linking halkeriids to mollusks.
Brachiozoa?: Cohen et al., 1998 and Cohen and Weydmann, 2005 indicated that phoronids are a valveless form of brachiopod. More recently, however, Hausdorf et al. 2010 find Phoronida to be the sister taxon of a monophyletic Brachiopoda. They name the Brachiopoda + Phoronida clade "Brachiozoa."
Always encased inside brachial and pedicle valves (except for pedicle)
Always benthic marine
Possesses
Nephridia for waste excretion
Closed circulatory system with a distinct heart but which doesn't seem to be involved in gas transport. (In contrast to Phoronida.)
Coelomic fluid with the oxygen-binding protein hemerethryn.
"Mantle tissue" that secretes the valves. (Not homologous to mollusk mantle.)
Simple nervous system coordinated by one or two ganglia near the base of the lophophore. Sensory neurons concentrated near anterior portion of the valves.
Living brachiopods are sensitive to sudden changes in illumination, and avoid light generally, preferring cryptic environments.
Brachiopod flesh is bad-tasting and avoided by potential predators.
Refer to lab for details of valve morphology.
Antarctic terebratulid brachiopods from BBC Nature
Brachiopod ecology: Although brachiopods encompass much diversity, certain generalizations are valid throughout.
All are shallow marine benthic
They typically require hard substrate. Those without pedicles (E.G. concavo-convex Chonetes) lay on soft substrate. Some were cemented to the substrate.
The utility of such strategies depended on the energy of the environment.
Moderate energy: E.G. a reef environment below low tide line. A direct pedicle or spiny attachment to a rigid substrate or host.
Low energy only: E.G. a sandy bottom below wave base. Rafting on soft sediment. (Even moderate energy here would bury the rafter.)
High energy: E.G. Intertidal zone. Here, sessile creatures must be firmly attached to a hard substrate, as in craniiform brachiopod cemented to strophomenid brachiopod.
Positioning and attachment: Position has two crucial aspects:
Location
Orientation
Typically, sessile organisms have to attach to rigid substrates. This limits their feeding to within a few cm. of the bottom, and excludes them from soft (sandy or muddy) substrates. E.G.: brachiopods typically attach to the substrate using a very short fleshy stalk, the pedicle.
Orientation: Using the pedicle, it can adjust its orientation to optimize suspension feeding. Its feeding, however, is limited to within 2 - 3 cm of the substrate, and the substrate must be rigid.
Location: Some brachiopods overcame this limitation by attaching to larger suspension-feeding organisms like stalked echinoderms, either with their pedicle (in which case they could still control their orientation somewhat) or using spines on their valves (in which case they depended on the substrate-host to orient them properly).
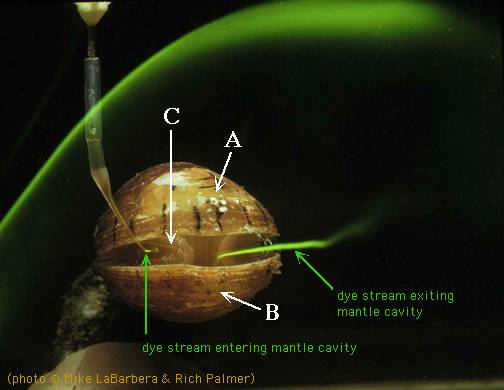
Brachiopods retain strong bilateral symmetry, even internally. Food particles are filtered from water brought inside the valves and mantle cavity by mucus secreted by the the lophophore tentacles. Intercepted particles are moved to the mouth by ciliary action. Upper and lower valves control the flow of the ciliary current across the lophophore, eliminating the need to be able to capture food coming from any direction.
Crenulations: In many bivalved organisms, most notably brachiopods, the commissure at which the valves meet forms a series of zig-zag crenulations. Functions include:
Strength: It is much more difficult for the valves of a crenulated shell to be twisted apart.
Major clades: Biostratigraphers and other nuts-and-bolts employers of practical paleontology have long employed a traditional Linnean taxonomy of brachiopods, that has been unsettled by the rise of Phylogenetic Systematics. The application of the latter has resulted in the elevation of previously obscure lower-order groups to prominence and the dismissal of time-honored groups as paraphyletic. Here, we attempt a compromise: a survey of the phylogenetic taxonomy of the major traditional Linnean groups based primarily on Carlson, 2007. For a more detailed phylogeny see 3D Brachiopod Images at UC Davis.
In traditional taxonomy brachiopods were divided between "articulates" - those possessing tooth-in-socket hinged articulations between valves, and "inarticulates" - those lacking such articulations that rely on soft tissue to control valves. The application of phylogenetic methods, in contrast, reveals three major clades:
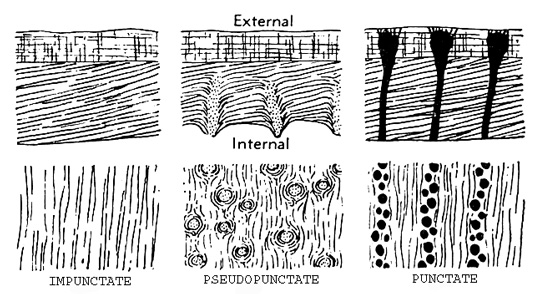
Valve histology: In cross-section, rhynchonelliform valves display obliquely layered inner layers of calcite overlain by low-angle lamellae. These can be:
Impunctate: if there is no suggestion of perforations
Interarea: The region between the valve's beak and hinge.
The pedicle foramen may take the form of a triangular delthyrium. This may be partly sealed by a distinct midline plate - the deltidium or by paired deltidial plates.
Pedicle variously developed, but always dead horny tissue, not invaded by the coelom.
Anus lacking. Gut tube is blind ended, with waste periodically being expelled through mouth. (In stark contrast to lingulids, which have a proper gut tube.) Note: The lophophorate filtration system normally rejects indigestible particles prior to ingestion, so the elimination of feces is not a big problem for these animals.
Epifaunal, with a wide variety of habits and substrate preferences, some of which change during ontogeny.
Rhynchonelliform systematics: A complex topic. Your text provides a detailed list of currently acknowledged monophyletic rhynchonelliform taxa. Here, we present seven groups that are heavily cited in traditional century literature. In some cases, their monophyly is not certain:
Plaesiomys subquadrata
"Orthida": (C - P. Abundant in C, O) Morphologically the most ancestral of articulates. Possibly paraphyletic.
Characteristics:
Distinct interarea with open delthyrium
Strophic
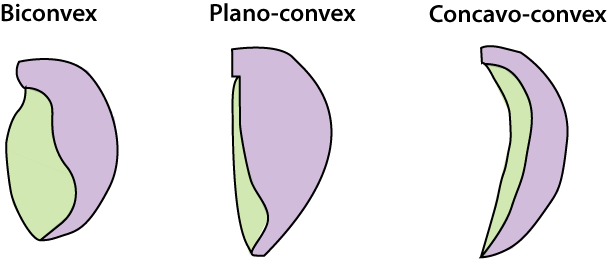
biconvex
Lacking elaborate brachidium
Raphinsequina sp.
Strophomenida: (O - Carb. Most abundant in Ordovician)
Characteristics:
Strophic with D-shaped profile.
Often lacking pedicle and colonizing soft substrates.
pseudopunctate
Frequently concavo-convex - an adaptation to "floating" on soft substrates.
Terbratulida: (D - Rec.) Never terribly abundant, but the most common post-Paleozoic brachiopods.
Characteristics:
Astrophic
Smooth biconvex valves
Looping brachidium
Their characteristic shape gives brachiopods the common name "lamp-shell."
Evolutionary trends: The relative abundance of different groups of these common fossils imparts a distinct character to shallow marine deposits of different systems. Thus even a geologist wit no interest in brachiopods, per se, should learn to identify them:
Ordovician-Devonian: Proliferation of rhynchonelliform brachiopods into deeper water, carbonate, and soft substrate environments. Strophic, D-shaped types such as strophomenids and orthids predominate. Severe reductions during Ordovician mass extinction with recovery during Silurian.
Carboniferous-Permian: Following the decline of stromatoporoids during the Late Devonian extinctions, brachiopods emerged in the Late Paleozoic as the major encrusting suspension-feeders of reef environments, but became less dominant on soft substrates:
Proliferation of encrusters, including odd-ball productids with specializations for cementation to hard substrate (E.g. Prorichthofenia, Leptodus - right).
This interval also saw the emergence of rhynchonellids and spiriferids as major components of the fauna.
Post-Paleozoic: The Permian extinction all but eliminated the more basal rhynchonelliformes, and the Triassic extinction finished off the survivors, leaving linguliforms, craniiforms, rhynchonellids, and terebratulids with essentially modern ecologies as marginal inhabitants of cryptic reef environments.
Brachiopod origins:
Xianshanella haikouensis
The brachiopod stem:
Tommotian stem brachiopods?: Members of the earliest Cambrian small-shelly fauna, including Micrina were proposed by Holmer et al., 2008 as near the ancestry of brachiopods.
Sutton et al., 2010 described Drakozoon kalumon (Silurian), a soft-bodied lophophore-bearing creature that might represent an intermediate grade between phoronids and brachiopods. Unlike phoronids, it attaches to the substrate by a broad attachment, but unlike brachiopods, has no mineralized valves - only a broad soft-tissue "hood."
Sun et al., 2018 described Pedunculotheca diania (Cambrian), an obvious member of the obscure Cambrian taxon Hyolitha with a peduncle! Apparently hyoliths are stem brachiopods.
Xianshanella haikouensis
Because creatures like Lingula show up early it is tempting to regard them as the ancestral brachiopod morphotype, especially given their mobility. Key Cambrian fossils seem to refute this:
Xianshanella haikouensis: Zhang et al. 2006. From Chengjiang, this early lingulid had a long pedicle but lived attached to hard substrates. (right)
Oldest crown brachiopod: This honor goes to Askepasma saproconcha from the Tommotian of Australia (Topper et al., 2013). Askepasma is a paterinate linguliform with a phosphatic shell, however it displays rhynchonelliform characters such as interareas, delthyria and notothyria, and proper diductor muscles. Could the absence of these features in other linguliforms be a reversal?
We are left with a picture of an attached ancestral brachiopod with an articulate but phosphatic shell. We await the key fossils.
For an outstanding review of brachiopod evolution, see Carlson, 2016
But what does any of this tell us about everyone's favorite group?
The taxonomic history of this group is complex. Nineteenth century workers first applied "Bryozoa" to the same critters that bear it today. By the turn of the 20th century, however, the discovery of entoprocts had complicated matters such that workers recognized two classes of "bryozoans:"
Ectoprocta, (the original "Bryozoa") whose anuses were outside the circle of their lophophores,
Entoprocta, whose anuses were inside this circle.
During the 20th century, these were recognized as being only distantly related. With Entoprocta removed, "Ectoprocta" became a junior synonym of Bryozoa. Today some workers continue to use Ectoprocta, however.
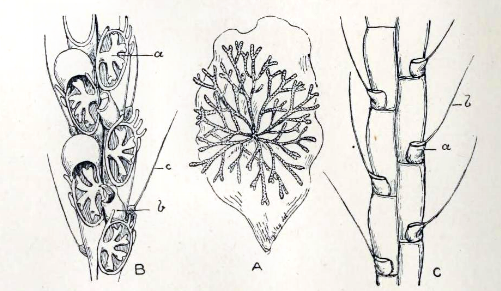
Colony (zoarium) begins with an initial zooid, the ancestrula, which buds off clones to form the remainder of the colony. (But a soft tissue connection persists.)
Individual zooids connected by funiculus and stolon through which nutrients are shared.
Lophophore is protracted when it "inflated" by coelomic fluid when circumferential muscles of body wall contract. It is retracted by retractor muscles. (In contrast, the lophophore tentacles of entoprocts are solid.)
Small enough to handle gas-exchange by simple diffusion.
Lack nephridia (organs for excretion of nitrogenous wastes) These, thus, accumulate in tissues.
Great powers of regeneration. Occasionally, all cells of the zooid except for the body wall degenerate into a brown body that is ultimately ejected from the zooecium. The zooid then regenerates from the body wall. Possible functions:
Resting stage during stressful intervals
A radical means of ridding the body of insoluble wastes.
Reproduction of both zooids and colonies is often by budding. Individuals reproduce sexually, as well. Most zooids are hermaphroditic, although ovaries and testes are usually not active simultaneously.
Ova are retained in the zooecium
free-swimming sperm are snared by the lophophore and conveyed to the ovum.
The zygote escapes as it transforms into a planktonic larva.
Zooecial walls incomplete, allowing zooids freely to share coelomic fluid.
Freshwater
Can propagate through statoblasts - little capsules of cells enclosed in calcareous capsules that form along the funiculus. In times of stress, when the zooid dies, these disperse. When proper conditions return, they open and a new ancestrula regenerates.
encrusters with largely calcareous zoaria. Because the zooecium is rigid, hydrostatic protraction of the lophophore is facilitated by adaptations like:
Protractor muscles attach to a flexible frontal membrane inside of zooecium.
Zooecium contains a water-filled compensation sack that communicates with the exterior. This empties or fills to compensate for protraction or retraction of the lophophore.
Early Paleozoic: Trepostomata and Cryptostomata dominant in Ordovician, but sharply curtailed by Ordovician extinction, never regaining dominance and suffering again in Devonian and Permian events, with only some trepostomatids straggling into the Triassic.
Silurian - Devonian: Peak diversity for Cystoporata and Trepostomata.
Late Paleozoic: The age of Fenestrata, some of which are Carboniferous index fossils. These are extinguished by Permian event.
Late Mesozoic: Great proliferation of Cyclostomatida and Cheilostomata. All suffered in K-Pg extinction,
Cenozoic: The age of Cheilostomata. Cyclostomatida recovers from K - Pg extinction but never regains preeminence.
Lophophorate Phylogeny:
The Linnean "subkingdom" Lophophorata has been recognized as problematic since the early days of phylogenetic systematics, and remains unsettled.
Background:
Early applications of phylogenetic methods to the problem suggested that phoronids and brachiopods might be:
Interpretation of morphology was frustrated by ambiguities of homology, however. (E.G. the endodermal coelom of larval bryozoans is resorbed during metamorphosis and replaced by a new "coelom" resembling the three-part coelom of deuterostomes, except that it is derived from ectoderm.) Most recent clarity on this topic, therefore, comes from molecular phylogenies.
Numerous molecular studies support the monophyly of Lophotrochozoa, including (but not limited to) Trochozoa, Phoronida, Brachiopoda, and Bryozoa.
Bryozoans are classic "wild card" taxa, jumping around in otherwise stable phylogenies. They seem to be closer to Trochozoa than to Ecdysozoa or Deuterostomia in most molecular analyses. Alas, the bryozoan genome is as derived as its morphology, making it a difficult subject for molecular methods.
Bryozoa, not brachiopods, are the sister taxon to phoronids.
Brachiopods, are the sister taxon to (Bryozoa + Phoronida).
Thus, Lophophorata could actually be monophyletic!
The whole question of lophophorate phylogeny is unsettled. We seem to know only that:
They belong to a monophyletic Lophotrochozoa
We await:
Better fossils
Improved taxonomic sampling for molecular phylogenies
Better understanding of the complexities of the interpretation of molecular data.
Stay tuned.
Additional reading:
Carlson, S. J. 2007. Recent research on brachiopod evolution. Pp. H2878–H2900 in A. Williams et al. Brachiopoda 6 (revised), Supplement. Part H of P. A. Seldon, ed. Treatise on invertebrate paleontology. Geological Society of America, Boulder and University of Kansas, Lawrence.
Bernard L. Cohen, Agata Weydmann. 2005. Molecular evidence that phoronids are a subtaxon of brachiopods (Brachiopoda: Phoronata) and that genetic divergence of metazoan phyla began long before the early Cambrian. Organisms Diversity & Evolution 5(4): 253-273.
Peter Funch and Reinhardt Kristensen. 1995. Cycliophora is a new phylum with affinities to Entoprocta and Ectoprocta. Nature 378, 711 - 714.
Halanych KM, Bacheller JD, Aguinaldo AM, Liva SM, Hillis DM, Lake JA. 2010. Evidence from 18S ribosomal DNA that the lophophorates are protostome animals. Science 268(5210): 485.
Bernhard Hausdorf, Martin Helmkampf, Maximilian P. Nesnidal, Iris Bruchhaus. 2010. Phylogenetic relationships within the lophophorate lineages (Ectoprocta, Brachiopoda and Phoronida). Molecular Phylogenetics and Evolution 55(3): 1121-1127.
Martin Helmkampf, Iris Bruchhaus, Bernhard Hausdorf. 2008. Phylogenomic analyses of lophophorates (brachiopods, phoronids and bryozoans) confirm the Lophotrochozoa concept. Proceedings of the Royal Society B 275(1645).
Holmer L. E., Skovsted C. B., Brock G. A., Valentine J. L., Paterson J. R. 2008. The Early Cambrian tommotiid Micrina, a sessile bivalved stem group brachiopod. Biology Letters 4(6):724-8.
Kuem Hee Jang and Ui Wook Hwang. 2009. Complete mitochondrial genome of Bugula neritina (Bryozoa, Gymnolaemata, Cheilostomata): phylogenetic position of Bryozoaand phylogeny of lophophorates within the Lophotrochozoa. BioMed Central Genomics 10(167).
M. P. Nesnidal, M. Helmkampf, A. Meyer, A. Witek, I. Bruchhaus, I. Ebersberger, T. Hankeln, B. Lieb, T. H. Struck, B. Hausdorf. 2013. New phylogenomic data support the monophyly of Lophophorata and an Ectoproct-Phoronid clade and indicate that Polyzoa and Kryptrochozoa are caused by systematic bias. BioMed Central Evolutionary Biology 13(253).
Jordi Paps, Jaume Baguñá and Marta Riutort. 2009. Lophotrochozoa internal phylogeny: new insights from an up-to-date analysis of nuclear ribosomal genes. Proceedings of the Royal Society B 276: 1245-1254.
Haijing Sun, Martin R. Smith, Han Zeng, Fangchen Zhao, Guoxiang Li, Maoyan Zhu. 2018. Hyoliths with pedicles illuminate the origin of the brachiopod body plan. Proceedings of the Royal Society B 285 (1887).
M. D. Sutton, D. E. G. Briggs, David J. Siveter, Derek J. Siveter. 2010. A soft-bodied lophophorate from the Silurian of England. Biology Letters 7(1).
T. P. Topper, L. E. Holmer, C. B. Skovsted, G. A. Brock, U. Balthasar, C. M. Larsson, S. P. Stolk,
and D. A.T. Harper. 2013. The oldest brachiopods from the lower Cambrian
of South Australia. Acta Palaeontologica Polonica 58 (1):
93–109..
Zhifei Zhang, Lars E. Holmer, Christian B. Skovsted, Glenn A. Brock, Graham E. Budd, Dongjing Fu, Xingliang Zhang, Degan Shu, Jian Han, Jianni Liu, Haizhou Wang, Aodhan Butler, and Guoxiang Li. 2013. A sclerite-bearing stem group entoproct from the early Cambrian and its implications. Scientific Reports 3(1066)
Zhifei Zhang, Degan Shu, Jian Han, Jianni Liu. 2006. New Data on the rare Chengjiang (Lower Cambrian, South China) linguloid brachiopod Xianshanella haikouensis. Journal of Paleontology 80(2):203-211.





































{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
.jpg){kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}