Fall Semester 2020 Ecdysozoa I: Basal Panarthropoda
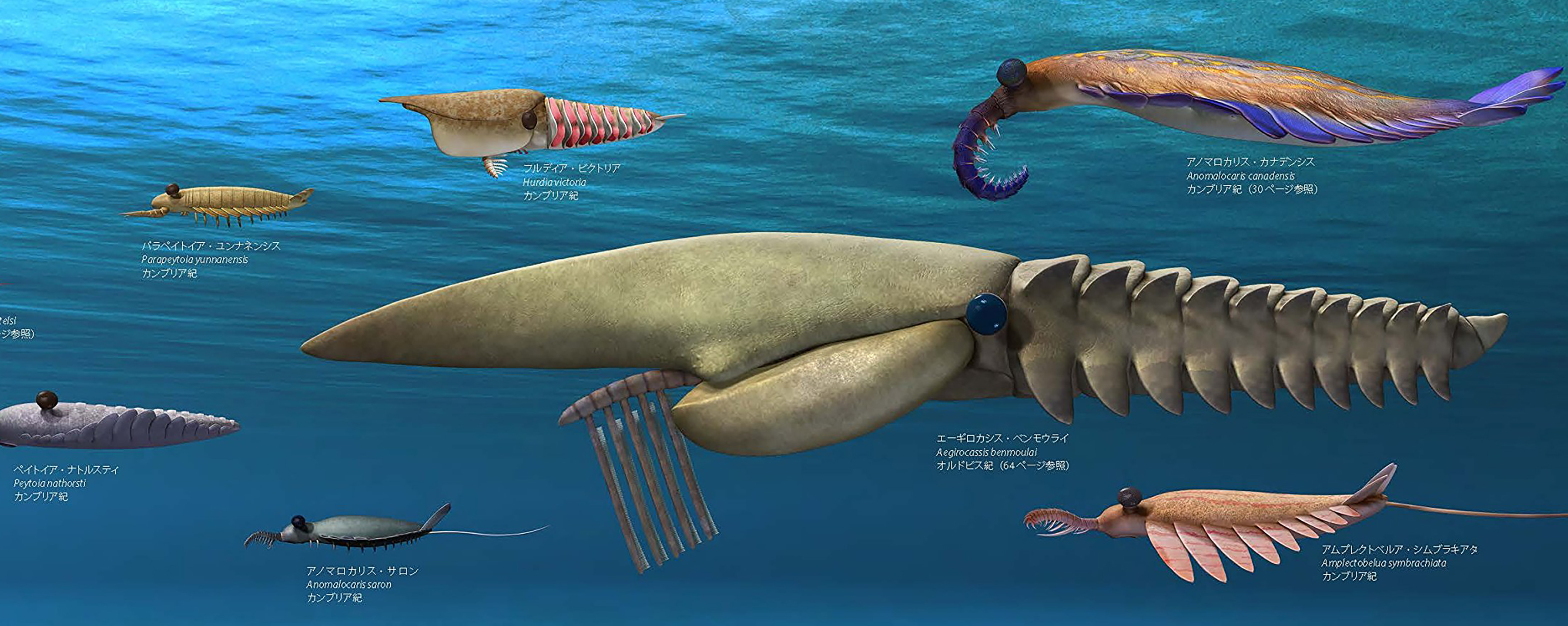
Cambrian radiodonts prefigure Arthropod diversity
Key Points:
•Ecdysozoa, sister taxon to Lophotrochozoa, contains Panarthropoda plus a variety of from-shaped invertebrates including Kinorhyncha, Priapulida, and Nematoda.
•Diagnosed by regular ecdysis and a terminal mouth lined with a circle of denticles.
•Panarthropoda adds distinct paired limbs with circles of stiffened cuticle and segmented bodies with repeating units.
•Living Panrthropods include Onychophora, Tardigrada, and Arthropoda.
•Panarthropods appear in the Early Cambrian. Members include Mureropodia, "lobopodan-grade" organisms, "Dinocarid-grade" organisms, Schinderhannes, and proper Arthropoda.
•Arthropoda is diagnosed by properly "arthrodized" cuticle, where cuticle segments meet in articulated joints - an apomorphy-based definition!.
•Euarthropoda - the crown group of living arthropods, includes Arachnamorpha and Mandibulata.
•The arthropod stem includes primitive bivalve arthropods, Fuxianhuiida, and arthropods of "megacheiran-grade.".
•Arachnomorpha contains Trilobita and Chelicerata. Trilobites represent the first very speciose flowering of arthropod diversity.
•Trilobites range throughout the Paleozoic, but have their peak diversity and abundance in the Ordovician.
•We review the anatomy of trilobites and the phylogenetic and ecological signal of anatomical variation.
"To a good approximation, all species are insects."
(Robert May.)
Ecdysozoa I: Basal Panarthropoda
Ecdysozoa: Organisms characterized by the regular shedding of an external cuticle and protostomous development.
Probable sister taxon to Spiralia (see review by Halanych, 2004.)
Burgess Shale priapulid Ottoia prolifica from Wikipedia
The monophyly of Ecdysozoa is well supported. It includes:
Limbs are moved in part by muscles and in part by coelomic hydrostatic skeleton.
Terrestrial onychophorans have no fossil record but for a single Late Carboniferous Mazon Creek fossil (Thompson and Jones, 1980). However, many marine fossils of the Cambrian resemble them superficially.
Tardigrada (Cambrian - Quaternary). AKA "water-bears." Cute. Miniaturized lobopodans of damp and aquatic habitats. These are minute and anatomically secondarily simplified. Feed on plant cells with piercing anterior mouth-parts. Only fossils known are from middle Cambrian Orsten Lagerstätte (Maas and Waloszek, 2011.)
Sally-lightfoot crab Grapsus grapsus
Arthropoda (Cambrian - Quaternary). The overwhelming majority of living ecdysozoan species. Where to start? All are characterized by a stiff cuticle that covers the body and forms distinct limb joints.
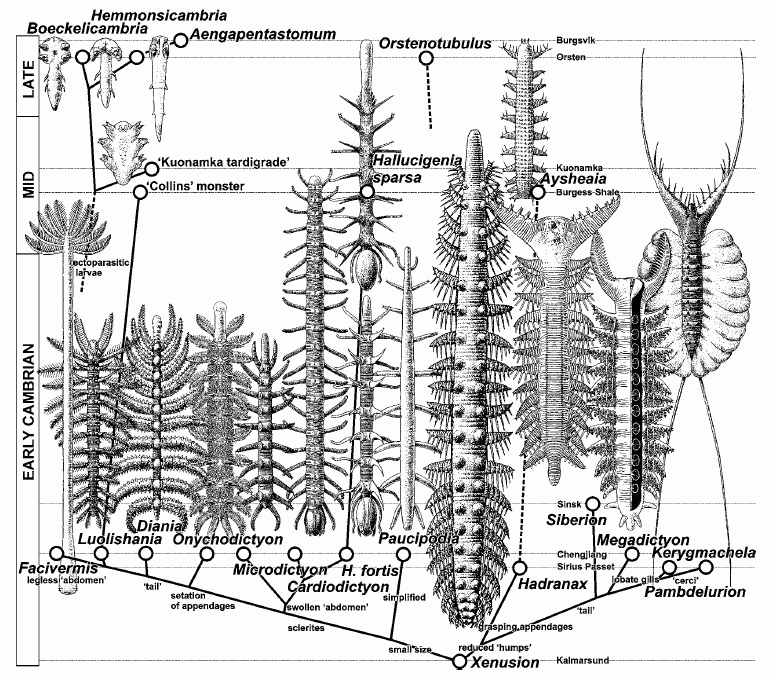
Fossil Panarthropoda: The Cambrian Chengjiang, Burgess Shale, and Sirius Passet provide a significant record of stem panarthropods. Alas, their literature tends to lack phylogenetic rigor and to conflate creatures that are superficially similar. One often hears group names that are definitely paraphyletic like:
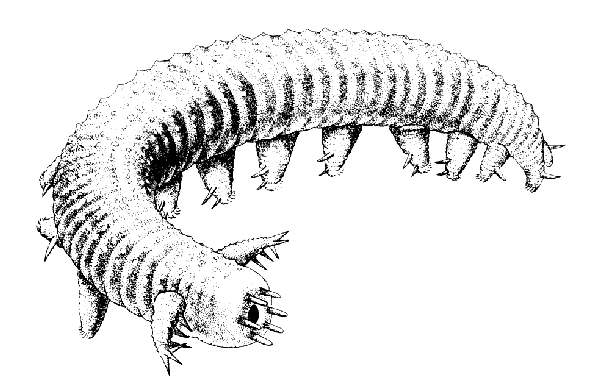
Lobopoda: Creatures broadly resembling marine onychophorans like the Chengjiang (Cambrian) Diania.
Dinocarida: Creatures with lateral swimming flaps and large raptorial (prey-capturing) anterior appendages like Anomalocaris.
We follow the phylogenetic result of Liu et al., 2011. Simplified at right. Our outgroup and starting point is Priaulida, whose members lack segmentation and limbs, and whose mouth consists of radially symmetrical parts that are protracted by hydrostatic action. As we climb the tree, be aware of:
Specialization of limbs
The position and orientation of the mouth
Evolution of specialized mouthparts
Evolution of sense organs
The origin and proliferation on the body of arthropod-style cuticle
Mureropodia apae (early Cambrian) An infaunal creature believably transitional between priapulids and panarthropods, with five pairs of flexible unspecialized limbs, and a proboscis with a terminal mouth.
Aysheaia pedunculata (middle Cambrian) The classic Burgess Shale "lobopodan." Sometimes called an ancestral marine onychophoran, but actually lacking many of their synapomorphies. Aysheaia occupies a basal position in recent analyses because of its lack of robust cuticle and specialized mouthparts. It does show the specialization of the first pair of limbs for something (?) like prey capture or as a sense organ.
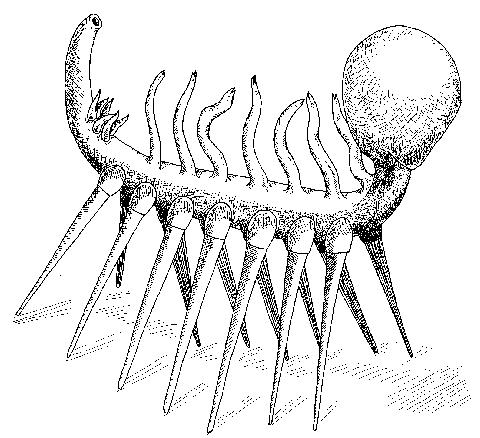
Stem onychophorans (Cambrian) Closer to Onychophora. Poorly understood prior to 2000. The first representatives to be studied inlclded Hallucigenia of the Cambrian Burgess Shale, a creature so poorly understood as initially to be misrepresented to the public as a stilt-walking worm. The discovery of less extreme armored forms like Microdyction (right) facilitated proper interpretation.
All appear to have specialized anterior appendages.
in a arrangement reminiscent of proper arthropod biramous appendages. Adding color to this similarity is the fact that in most taxa, these swimming appendages also seem to be house gas-exchange organs.
Like the previous taxa, they retain and elaborate on specialized appendages, sporting an anterior pair of "great appendages." On the opposite end, they possess abdominal cerci (sensory (?) appendages) homologous (?) to those of arthropods.
Interesting examples include:
Kerygmachela kierkegaardi: (early Cambrian from Sirius Passet) With small eyes and a terminal mouth. (~ 20 cm.)
A famous example is the five-eyed Opabinia regalis (middle Cambrian Burgess Shale). Unique in that its great appendages are fused into a single mid-line appendage.
They are derived in:
The presence of Compound eyes: composed of clusters of ommatidia (singular "ommatidium") - visual elements each with a lens and visual cells.
The position of the mouth beneath the head.
(Aside: In your instructors' opinions, the alleged Cambrian proto-cephalopod Nectocaris pteryx, (Smith and Caron, 2010) belongs somewhere in this region of the tree.)
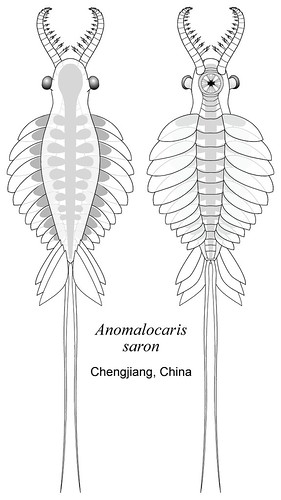
Radiodonta (Cambrian). (The derived "dinocarids." Commonly known as "anomalocarida" or "anomalocaridida," we embrace the phylogenetically defined (Liu et al., 2011) name of this monophyletic group.) The giant (0.5 m) top predators of the Cambrian. Morphologically similar to less derived "dinocarids," but:
Ecology: Radiodonts, as apex predators, were much larger than other Cambrian animals. We have begun to notice a surprising diversity of function in their great appendages. Most are predators who used these limbs to grab prey, but others may have been scavengers (Hurdia) or, in the case of Aegirocassis, even ram suspension feeders like baleen whales. Like whales, they dwarfed other creatures in their environment.
Schinderhannes bartelsi:Kühl et al., 2009 reported a small (10 cm) radiodont-like animal Schinderhannes bartelsi (right) from the Devonian of the Hunsrück lagerstätte. Like radiodonts and "dinocarids," it retained impressive great appendages and a circular toothy mouth. In other respects, it is closer to living arthropods:
The radiodont swimming appendages are replaced by proper jointed biramous appendages
The dorsal body cuticle is divided into distinct tergites.
A telson - posterior spine - is present.
The "mustache" is a unique feature of Schinderhannes. Is it a proper arthropod? Opinion is divided but it is certainly close.
Records and rumors: Reliably identifiable fossils of panarthropods date no farther back than the Early Cambrian. Nevertheless, some researchers claim to recognize them in the zoological Rorschach-test of the Ediacaran. Putative Ediacaran panarthropods might include:
Parvancorina: (Right) Small shield-shaped creatures with body sculpture reminiscent of the cephala and pleural and axial lobes of trilobites. The case for Parvancorina draws strength from its similarity to the Burgess Shale trilobitomorph Skania fragilis.
Spriggina floundersi: An Ediacaran animal. Rangeomorph frond? Annelid? Panarthropod? Each hypothesis has supporters.
Alas, the preservation of Ediacaran fossils typically precludes the identification of potential structures like jointed limbs on tiny Parvancorina, reducing our attempts to identify their affinities to paleontological pareidolia.
Arthropoda:
(Cambrian - Quaternary) Arthropods ("Euarthropoda" of some authors, used herein to refer to the crown group of Arthropoda) with a proper arthrodized (sclerotized and jointed) cuticle covering the body have a copious fossil record, as even unmineralized cuticle is a "tough tissue," facilitating copious preservation in Konzervatlagerstätten. Some groups - Especially trilobites, mineralize their cuticle in addition. During the last twenty years, their phylogenetic pattern has began to come into focus, as well.
Each body segment is associated with a pair of limbs or their derivatives except for:
The acron: the first (pre-oral) segment
The telson: the final abdominal segment (which often takes the form of a spine.)
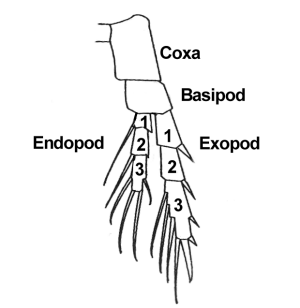
Ancestrally limbs seem to be biramous (right): one branch, the endopod (ventral), for locomotion, the other, the exopod, (dorsal) branch
being specialized as a gas exchange structure (gill).
Various body segments of various derived forms lose either the dorsal or ventral ramus to become uniramous
Specialized feeding and sensory appendages are modified from walking limbs.
Body segmentation is complete.
Body of primitive forms is homonomous (all body segments are very similar), but in
vast majority there is a significant degree of tagmosis (AKA "tagmatization") - specialization and fusion of body segments and their associated limbs. Patterns of tagmosis, the grouping of segments into tagmata - functional regions of the body - are very significant to arthropod systematics, starting with the assessment of the homologies of segments and their limbs.
Nitrogenous waste eliminated by malphigian tubules
Heart with pericardium and ostia (sing. ostium). Open circulatory system with large haemocoel (coelomic cavity specialized for the transmission of circulatory fluid).
Highly cephalized: Paired nerve cords run ventrally to the gut tube. For each segment, each cord has a ganglion from which the segment is innervated. Each pair of ganglia is connected by a commissure. The first three meet in a commissure in the head, dorsal to the esophagus, forming a brain.
The living euarthropod groups:
Nephila edulis
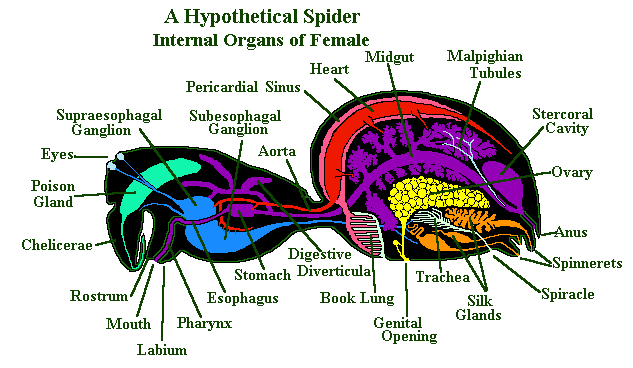
Chelicerata: (Cambrian - Quaternary) Including
Xiphosura - horseshoe crabs
Arachnida - spiders (right), scorpions, ticks, mites, and many more.
Pycnogonida - "sea-spiders" (but not really)
Extinct members including Eurypterida - "sea scorpions"
Narceus sp.
Myriopoda: (Silurian - Quaternary) Including
Diplopoda - millipedes (right)
Chilopoda - centipedes
Blue crab - Callinectes sapidus
Pancrustacea: (Cambrian - Quaternary) Including
Hexapoda - Insects and their kin
A bewildering variety of traditional "crustaceans" (paraphyletic)
Arthropod Phylogeny
Molecular and morphological phylogenies of the last decade (especially the molecular analysis of Regier et al., 2010. have resolved the major patterns of the relationships of the living taxa, and major fossil ones. Among Euarthropoda, the arthropod crown group, two major groups are evident:
Arachnomorpha: Trilobites, Chelicerates, and their kin.
Mandibulata: Centipedes, crustaceans, insects, and their kin.
The major ongoing controversy concerns Pycnogonida - the "sea-spiders," recovered by some as basal chelicerates and by others as the sister taxon to the remaining euarthropods.
Things become more interesting when the fossil arthropods of the Cambrian Konzervatlagerstätten are considered. During the last three years, however, a picture has begun to emerge. A survey of "major" groups.
"Bivalve arthropods" (Cambrian). Not to be confused with bivalved members of Euarthropoda like ostrocods, this is a paraphyletic grade at the base of Arthropoda. Basal arthropods like Nereocaris exilis (right) possessed:
A thoracic shield hinged on the midline
Two lateral compound eyes plus a third rod-shaped midline eye.
Fuxianhuiida (Cambrian). Chengjiang forms including Fuxianhuia (right) with distinct segmented body cuticle. Has anterior stalked eyes, six pairs of head appendages, two - three pairs of simple jointed limbs per body segment. The trunk is divided into a head-shield, an anterior limb-bearing thorax, a posterior abdomen, and ends with a pointed telson.
Recently, fuxianhuiids have become the poster children of arthropod phylogeny, with spectacular specimens revealing details of the nervous (Ma et al., 2012) and vascular systems (Ma et al., 2014). These reveal that in Fuxianhuia, these systems resemble those of living arthropods. Specifically, the brain is similar to those of most living crustaceans and insects, and its eyes and antennae are innervated similarly by the first and second sets of brain ganglia respectively.
"Megacheira" or "Great-appendage arthropods": A striking array of Cambrian arthropods display large raptorial anterior appendages. Examples include Leanchoillia superlata (right) and Yohoia tenuis from the Burgess Shale. Their great appendages are currently thought to be homologous to the antennae of most arthropods and the chelicerae of chelicerates, but not to the forelimbs of anomalocarids. Their position in arthropod phylogeny is debated:
Kühl et al., 2009: indicate they these are a paraphyletic group closer to Cheliceramorpha than to Trilobitomorpha.
Interpolating these basal groups, we get this basic phylogeny. Now to explore.
Arachnomorpha:
Contains:
"Trilobitomorpha": (Cambrian - Permian) Trilobites and their relatives. Whether this group is mono- or paraphyletic is currently unclear. Trilobita is certainly monophyletic.
Cheliceramorpha: (Cambrian - Quaternary) Chelicerates (horseshoe crabs, sea scorpions, arachnids) and their relatives.
Extremely common fossil makers, particularly in the early Paleozoic
Range from early Cambrian, when they were major players in the "Cambrian explosion" to the Permo-Triassic Extinction. They experienced their peak diversity in the Ordovician and were sequentially reduced by extinction events at the end of the Ordovician and the Devonian. Only a single clade, Proteida, straggled through the late Paleozoic, only to be snuffed at the end Permian extinction.
Exoskeletons were highly calcified, and thus very well preserved in comparison to their relatives, who show up only at sites of exceptional preservation (E.G. Konzervatlagerstätten: Chengjiang, Sirius Passet, Burgess Shale.) These relatives include minor groups such as:
Naraoida: Similar to trilobites without calcification or segmentation of thorax
Tegopeltida: Similar to trilobites without distinct cephalon
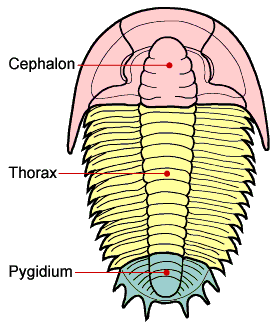
Trilobite tagmosis:
The trilobite body is divided into three regions:
Cephalon: (Head) with paired compound eyes, one pair of antennae )in front), and a pair of unspecialized biramous legs for each posterior segment
Thorax: One pair of biramous legs for each segment.
Pygidium: Posterior segments fused into a plate. Each segment with a pair of unspecialized biramous legs. In some cases a pair of antenna-like cerci are associated with the last segment.
Note: Some trilobites of the Early Cambrian (E.G. Olenellus) retain a telson, as well.
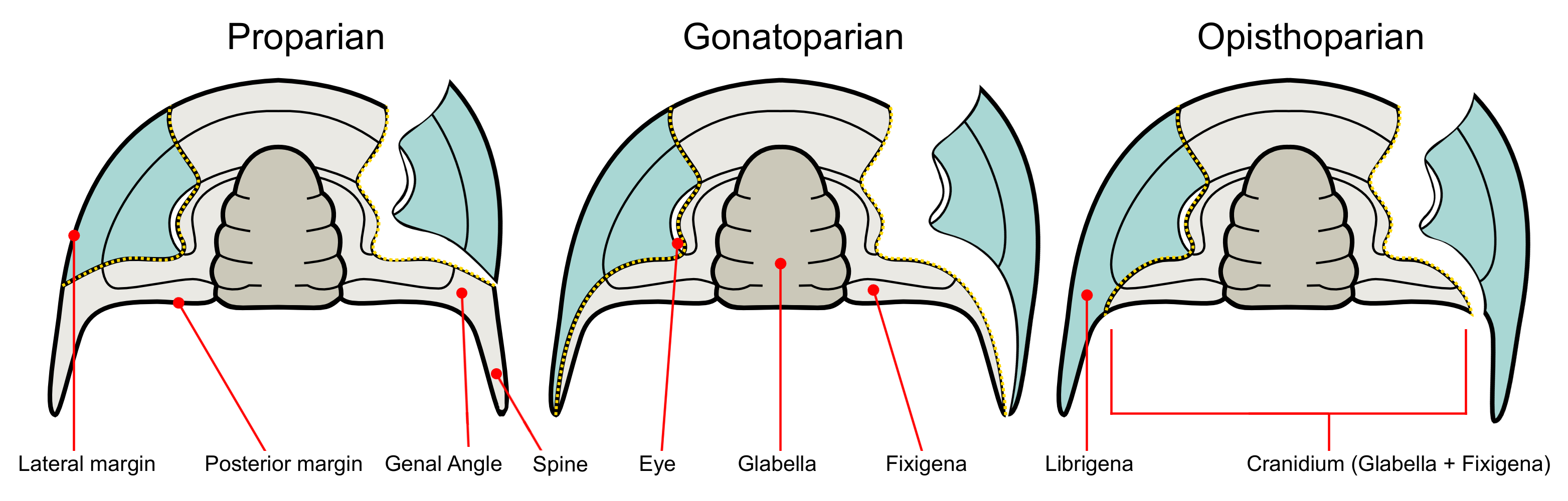
Facial sutures: The cephala of most trilobites display facial sutures - lines of weakness at which the cuticle of the cephalon breaks apart when the animal molts. In traditional taxonomy, their geometry has great significance. They are classified based on where the suture terminates posterolaterally with respect to the corner of the cephalon:
Proparian: sutures meet the lateral margin anterior to the corner.
Opisthoparian: (right) sutures meet the posterior margin of the cephalon.
Gonatoparian: sutures extend precisely to the corner of the cephalon.
Trilobite body divisions
Anatomy:
Antero-posteriorly into a cephalon (head, containing most of the viscera),
thorax (generaly with many segments, containing the limbs), and pygidium (fused "tail" segments)
Ventrally, the only calcified cuticle is in the hypostome, a plate that forms the floor of the mouth.
Medio-laterally into a central axial lobe (containing the nervous and digestive
systems) and a pair of lateral pleural lobes (overlying the spread of the limbs, including
their gills)
Trilobite alimentary canal
The mouth was ventral and opened into an expansion of the stomach that filled the forehead-like glabella.
Compound eyes were well developed. Lenses were single crystals of calcite with their crystalline axes aligned with the long axis of the eye. Two major and one minor type of eye are known:
Holochroal: Similar to the compound eye elements of insects. Lens elements are contiguous and focus onto a point. A single cuticle "cornea" covers the entire eye.
Schizochroal: (Right) Unique among animals to the phacopid trilobites. Lens elements are:
separated by thick cuticle
consist of two subunits with slightly different indices of refraction, creating an aplanatic correcting lens such as described by Huygens and Descartes
equipped with their own distinct cornea
Thus, each element creates a sharper image and transmits more light.
A third type, Abathochroal: Found in a handful of basal trilobites. Simple lens elements are separated by thin cuticle. A single cuticle "cornea" covers the entire eye.
Trilobite ecology: The diversity of eye morphology and degrees of visual acuity in trilobites prefigures their great range of ecological specializations. While most were benthic deposit feeders, some developed the ability to capture larger prey while others got up into the the water column. To scratch the surface:
Trilobite attacks prey
Very diverse feeding habits, including:
Predation/scavenging: This appears to be the ancestral state, present in some outgroups to trilobites such as Naraoia. Present in the largest forms. Presence indicated by spiny gnathobases and elongate, stout hypostomes.
Particle feeding: Most common forms (numerically and taxonomically), "elbowing" particulate food into their mouths. Indicated by modest gnathobase spines and reduced hypostomes.
Olenimorph trilobite
Chemosymbiotic forms:
Some forms, particularly of the clade Olenimorpha, found in deep water black shales seem poorly equipped to feed at all. Indeed, they resemble a trilobite imitating an ediacaran mat-animal, with a great many thoracic segments and very broad pleural lobes. Some speculation maintains that they were chemosymbiotic forms analogous to the pogonophoran worms of recent deep sea hydrothermal environments.
Pelagic planktonivores:
During the Ordovician, several groups gave rise to small nektonic plankton eaters. These are characterized by reduced cuticles, and enlarged eyes and limbs.
A particularly early and distinctive group, the Cambrian agnostids were, themselves, planktonic, experimenting with the trilobite version of the bivalve morphology found today in planktonic crustaceans. Some suggest that these are not actually trilobites. Owing to their highly derived, arguably paedomorphic, nature and our lack of detailed knowledge of the anatomy of other trilobites at similar developmental stages, this has been difficult to assess. Right now, opinion is divided.
Trilobite development
Trilobites being arthropods, we have good growth series for many of them, This enables us to identify agnostids as paedomorphic.
Trilobite systematics: Here we have a problem. Being well known for close to two centuries and principle index taxa for the early Paleozoic, their complex, traditional Linnean taxonomy is studied by a great many professional biostratigraphers. This includes some groups that are probably paraphyletic (See the Prothero text for a full review.) Consequently, few phylogenetic investigations have been conducted of the relationships within Trilobita. (See review by Lieberman and Karin, 2010.) This is bad because trilobites have been used in the study of evolutionary patterns and processes despite our not really knowing their real evolutionary pattern.
To date, most rigorous phylogenetic analyses have aimed at:
The position of trilobites in the broader arthropod context
We await a full-on cladistic analysis encompassing the full range of trilobite diversity. Indeed, we don't really know which trilobites are the basal group. Two candidates are:
Olenellida: Whose members retain a telson and lack facial sutures.
Agnostida: Whose tiny members lack even a hypostome. But note: Legg et al., 2013 place agnostids well outside of Trilobita!
Resolutions of such issues will influence our view of the ancestral ecology of trilobites.
Gabriele Kühl, Derek E. G. Briggs, Jes Rust. 2009. A Great-Appendage Arthropod with a Radial Mouth from the Lower Devonian Hunsrźck Slate, Germany. Science 323(5915): 771-773.
David A. Legg, Mark D. Sutton, Gregory D. Edgecombe, Jean-Bernard Caron. 2012. Cambrian bivalved arthropod reveals origin of arthrodization. Proceedings of the Royal Society B 279(1748).
David A. Legg, Mark D. Sutton & Gregory D. Edgecombe. 2013. Arthropod fossil data increase congruence of morphological and molecular phylogenies. Nature Communications 4(2485).
Bruce S. Lieberman, Talia S. Karim. 2012. Tracing the trilobite tree from the root to the tips: A model marriage of fossils
and phylogeny. Arthropod Structure & Development 39: 111-123.
Jianni Liu, Michael Steiner, Jason A. Dunlop, Helmut Keupp, Degan Shu, Qiang Ou, Jian Han, Zhifei Zhang & Xingliang Zhang. 2011. An armoured Cambrian lobopodian from China with arthropod-like appendage". Nature 470 (7335): 526-530.
Xiaoya Ma, Peiyun Cong, Xianguang Hou, Gregory D. Edgecombe, and Nicholas J. Strausfeld. 2014. An exceptionally preserved arthropod cardiovascular system from the early Cambrian. Nature Communications 5(3560).
Xiaoya Ma, Xianguang Hou, Gregory D. Edgecombe, and Nicholas J. Strausfeld. 2012. Complex brain and optic lobes in an early Cambrian arthropod. Nature 490, 258-261.
Jerome C. Regier, Jeffrey W. Shultz, Andreas Zwick, April Hussey, Bernard Ball, Regina Wetzer, Joel W. Martin, and Clifford W. Cunningham. 2010. Arthropod relationships revealed by phylogenomic analysis of nuclear protein-coding sequences. Nature 463, 1079-1083.
Derek J. Siveter, Derek E. G. Briggs, David J. Siveter, Mark D. Sutton, David Legg, Sarah Joomun. 2014. A Silurian short-great-appendage arthropod. Proceedings of the Royal Society B 281(1778).
Martin R. Smith and Jean-Bernard Caron. 2010. Primitive soft-bodied cephalopods from the Cambrian. Nature 465, 469Đ472.
Ida Thompson and Douglas S. Jones. 1980. A possible onychophoran from the Middle Pennsylvanian Mazon Creek beds of northern Illinois. Journal of Paleontology 54(3): 588-596.






























{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}