

.jpg)
Out of the mist, one hundred yards away, came Tyrannosaurus Rex.
"It," whispered Eckels. "It......"
"Sh!"
It came on great oiled, resilient, striding legs. It towered thirty feet above half of the trees, a great evil god, folding its delicate watchmaker's claws close to its oily reptilian chest. Each lower leg was a piston, a thousand pounds of white bone, sunk in thick ropes of muscle, sheathed over in a gleam of pebbled skin like the mail of a terrible warrior. Each thigh was a ton of meat, ivory, and steel mesh. And from the great breathing cage of the upper body those two delicate arms dangled out front, arms with hands which might pick up and examine men like toys, while the snake neck coiled. And the head itself, a ton of sculptured stone, lifted easily upon the sky. Its mouth gaped, exposing a fence of teeth like daggers. Its eyes rolled, ostrich eggs, empty of all expression save hunger. It closed its mouth in a death grin. It ran, its pelvic bones crushing aside trees and bushes, its taloned feet clawing damp earth, leaving prints six inches deep wherever it settled its weight.
It ran with a gliding ballet step, far too poised and balanced for its ten tons. It moved into a sunlit area warily, its beautifully reptilian hands feeling the air.
"Why, why," Eckels twitched his mouth. "It could reach up and grab the moon."
-- from Ray Bradbury's 1952 short story "A Sound of Thunder"
BIG QUESTION: What can we discover about the locomotion, predation, and ecology of T. rex and its kin?
Discovery of the "Model Dinosaur"
Easily the most famous species of Mesozoic dinosaur, Tyrannosaurus rex has been featured in popular culture in books, movies, comics, toys, video games, and so forth since its discovery in 1905. But even more than that: it is the most studied of the non-avian dinosaurs. Essentially any form of paleontological analysis that can be conducted on dinosaurs is done to T. rex first (and sometimes, frustratingly, ONLY to T. rex, so that no actual significant comparison to other species can be made!) Why did this species, and its close relative, become the "model dinosaur"?
In part it is surely that it is a highly charismatic animal: among the very largest predators ever to stride the land, spectacularly different than any comparable living creature (an orca-sized flesh eater walking on bird legs). But there are pragmatic reasons for it being the focus of study. Unlike many giant dinosaurs it is known from multiple well-preserved partial skeletons documenting nearly every (but not quite all!) bone in the body. It is known from multiple growth stages, allowing ontogentic analyses to be done. Enough specimens are known that there are numerous histological samples, digital models, and CT scans from multiple specimens. And given its superlative size it allows paleontologists (and others: it is the species of dinosaur on which non-specialists are most likely to publish on!) to test the limits of such issues as locomotion, bite forces, population size, and so forth.
We have not always known about Tyrannosaurus and its relatives. Some isolated bones and teeth had been found in the Rocky Mountain West as early as the 1870s, but beyond documenting a gigantic carnivorous dinosaur little could be said about its biology. Other large theropods were known during the 19th Century: Megalosaurus of Great Britain and Dryptosaurus of New Jersey, both known from partial specimens; and Allosaurus and Ceratosaurus from relatively complete skeletons from the American West. These were all in the 6-8 m range: much bigger than modern terrestrial predators.
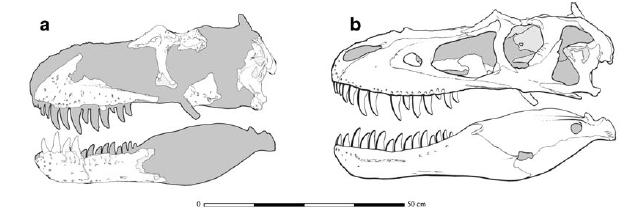
Expeditions by the Carnegie Museum of Natural History (in Pittsburgh) and especially the American Museum of Natural History (in New York) to Montana and Wyoming at the turn of the century began to find the first good fossils of a truly giant predator from the very end of the Cretaceous. A pair of skeletons (found in 1900 and 1905 by AMNH collector Barnum Brown) helped to describe this new species, which Brown's boss Henry Fairfield Osborn (whom we met before in mass extinctions, and whom we will meet again with regards to human origins) gave the most famous and memorable name in paleontology: Tyrannosaurus rex, "King of the Tyrant Lizards". At 12 m or more Tyrannosaurus dwarfed the previously-known theropods. Within a decade an even more complete skeleton with a complete skull made Tyrannosaurus the best understood carnivorous dinosaur. (Additionally, decades later this new specimen became literally iconic as it was featured in silhouette in the logo of a multi-billion dollar film franchise.)
However, Tyrannosaurus rex was not alone. In the same paper in which T. rex was first described, a slightly older and slightly smaller relative Albertosaurus sarcophagus was also named (based on a pair of skulls initially considered a new Canadian species of Dryptosaurus). Over the rest of the 20th (and on into the 21st) Century additional genera and species were added to this group, termed Tyrannosauridae. The tyrannosaurids shared a number of specialized features: fused nasal bones on the snout; incisor-like teeth in the front of the jaws (rare for dinosaurs); thickened teeth in the rest of the jaw (rather than the typical steak-knife teeth of most carnivorous dinosaurs); proportionately large heads with (for dinosaurs) proportionately large brains; binocular vision; reduced forelimbs with only two functioning fingers; enlarged hip muscles; proportionately long and slender legs with a specialized shock-absorbing foot (the arctometatarsus; and, of course, large body size.
Tyrannosaurids were limited in time and space. They are only present in the last 20 million years or so of the Late Cretaceous, and found only in eastern and central Asia and in western North America (the region called Laramidia): collectively, these regions (connected at times over a Siberian-Alaskan land bridge) is called "Asiamerica". Tyrannosaurids were not the only dinosaurs limited to this region: horned ceratopsids, domeheaded pachycephalosaurs, and club-tailed ankylosaurids were Asiamerican endemics, and this region was where the main diversity of ostrich-shaped ornithomimids and deinocheirids and duckbilled hadrosaurids were found. In Appalachia (eastern North America) the dryptosaurid relatives of tyrannosaurids were present alongside hadrosaurids and others, while in Europe and the southern continents entirely different groups of dinosaurs dominated.
If tyrannosaurids lived only at the end of the Cretaceous, where did they come from? Osborn favored a model where they were the direct descendants of earlier giant theropods (which I once termed the "super-carnosaur" model), while Brown and others considered them to be descendants of small predatory dinosaurs grown huge convergently with Allosaurus and its kin (what I called the "tyrannoraptor" hypothesis). The first detailed cladistic studies which considered the topic (including my own) supported the "tyrannoraptor" hypothesis, but clear evidence of pre-Late Cretaceous taxa closer to tyrannosaurids than to other Late Cretaceous groups was still lacking.
But throughout the last two decades numerous fossils of Jurassic and Cretaceous theropods on the tyrannosaurid lineage have been found. These are collectively the Tyrannosauroidea.
The oldest and most primitive are the Proceratosauridae. The oldest known are Middle Jurassic English Proceratosaurus and equally old Siberian Kileskus. Slightly younger (and far more completely known) is Guanlong of the early Late Jurassic of China: a 3 m or longer crested proceratosaurid. Like other early carnivorous dinosaurs, the arms were fairly long. Long arms with tridactyl hands were likely found in all proceratosaurids, but we do not yet have the arms for Proceratosaurus, Kileskus, or Early Cretaceous (and possibly 8 m long!) Sinotyrannus, one of the youngest known proceratosaurids. Of similar age (and possibly the same genus) is Yutyrannus of Early Cretaceous of China. Known from three nearly-complete fossils, this is a 9 m long predator. It still retained the tridactyl hand with long claws of typical theropods. Because they were preserved in fine-grained ash-based sediments, the remains of long protofeathers were found over its body, indicating that even giant theropods had a fuzzy coat!
Dilong of the Early Cretaceous of China represents the next phase of tyrannosauroid evolution, the Pantyrannosauria. It was the first tyrannosauroid discovered with protofeathers. At 1.5 m length, it still indicated that some basal tyrannosauroids were small members of the predatory community.
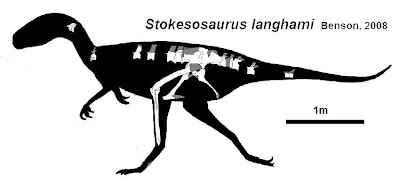
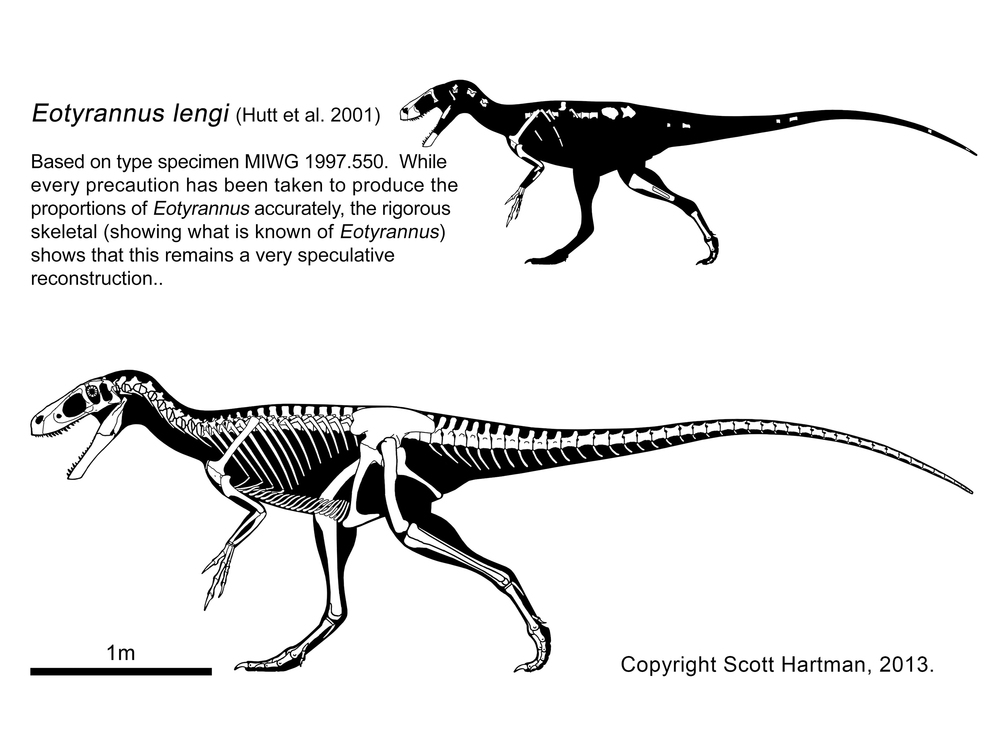
More robustly-built are the Stokesosauridae, including Jurassic tyrannosauroids such as Stokesaurus of North America, and Juratyrant (formerly considered a species of Stokesosaurus) of Europe, and Aviatyrannis of both. A later stokesosaurid is Early Cretaceous Eotyrannus of Europe, with an adult size of possibly 4.5 m or more. It was dwarfed by other theropods in its community: the carnosaur Neovenator and the spinosaurid Baryonyx. (A new analysis suggests that stokesosaurids actually branched off below the Proceratosauridae, and that Eotyrannus was actually a more derived tyrannosauroid closer to Xiongguanlong and later forms.)
Xiongguanlong of late Early Cretaceous China and Timurlengia of early Late Cretaceous Uzbekistan show an increase in body size and the development of increased bite forces.
Two small-bodied tyrannosauroids with arctometatarsi are known from the early Late Cretaceous of western North America: incompletely known Moros and more completely known Suskityrannus. Both lived in environments when other forms of theropods were the dominant apex predators. While the arms of proceratosaurids and stokessaurids were primitively long, those in Suskityrannus show the later tyrannosauroid trend of reduced arm length. Similarly, these forms show elongation of the distal hindlimb and the development of the arctometatarsus, traits shared with their more derived relatives.
The more derived Eutyrannosauria increase size again. One branch of eutyrannosaurs is the eastern North American 6 m or longer slender-footed Dryptosauridae, best-known from Dryptosaurus. In Dryptosaurus the arm is very short but has a very large claw. It is confirmed as having an arctometatarsus. As with more derived tyrannosauroids, the distal hindlimbs (tibia, metatarsi) are elongated: an indication of cursorial (running) ability. Another Eastern North American eutyrannosaur is Appalachiosaurus of the Late Cretaceous of eastern North America. About the same age is deep-skulled Bistahieversor of the Late Cretaceous of the American Southwest. But the best understood and most derived eutyrannosaurs were Tyrannosauridae proper.
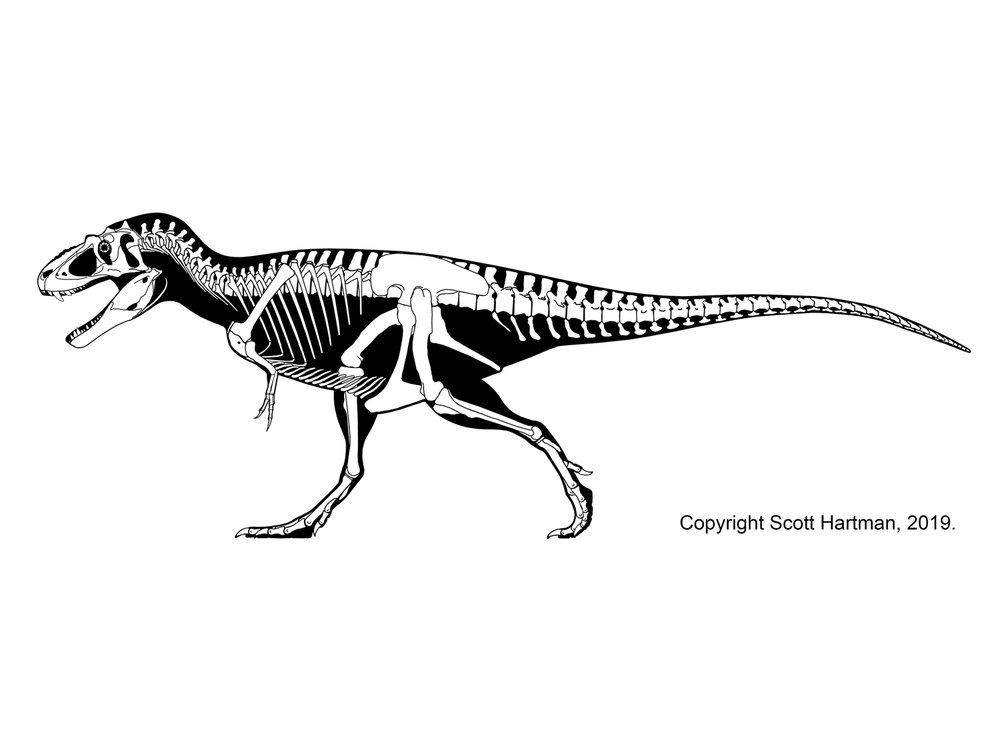
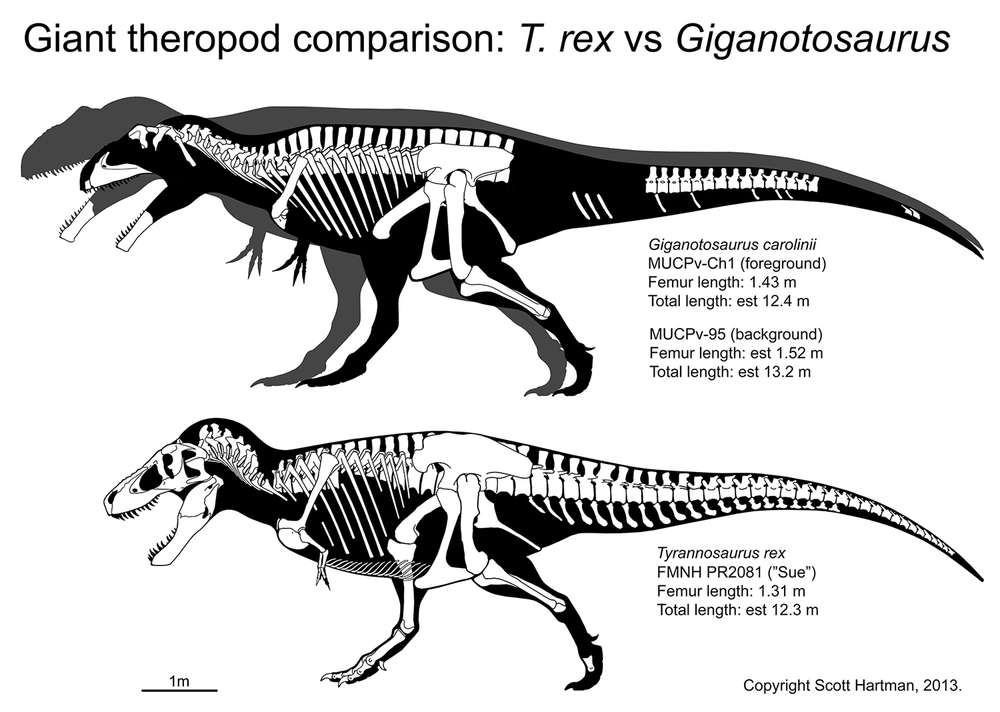
Tyrannosaurids include the relatively slender Albertosaurus and Gorgosaurus of western North America; slender long-snouted Alioramus and Qianzhousaurus of Asia; and more heavily built Lythronax, Daspletosaurus, and Teratophoneus of western North America and Tarbosaurus and Zhuchengtyrannus of Asia; and giant 13 m long, 8-10 ton Tyrannosaurus of western North America. Tyrannosaurus rivals the biggest carcharodontosaurs and spinosaurids in mass.
Tyrannosaurids are the largest theropod dinosaurs for which we have relatively complete remains. The mid-Cretaceous Spinosaurus was probably longer and may have grown as heavy, and the last of the giant allosaurs such as Tyrannotitan, Giganotosaurus, and Mapusaurus may have rivaled the biggest Tyrannosaurus in mass, but none of these are known from complete individuals. In contrast, at least two specimens 50% complete or more document individuals of T. rex greater than 8 tonnes. Its Asian cousins Zhuchengtyrannus and Tarbosaurus were 5 tonnes or more, and the others in the 2-3 tonne range.
With their great size came great power. The jaw muscle region of tyrannosaurids were expanded relative to other carnivores, and their teeth deeply rooted and thicker from side-to-side. Consequently they had much stronger bites than other theropod dinosaurs. Tyrannosaurid coprolites with pulverized dinosaur bone and bite marks from tyrannosaurids on hadrosaurids and ceratopsids document this force. Together, the deeply-rooted thick teeth and massive skull with powerful neck (and a solid palate to the roof of the mount), allowed tyrannosaurids to grasp onto their prey, breaking bone and holding onto flesh, and ripping and twisting off big chunks of meat. This "puncture-and-pull" method of feeding replaces the ancestral "bite-and-slice" method.
In contrast, the forelimbs are famously reduced. In adult Tyrannosaurus the tips of the claws can't even touch across the chest, and in Tarbosaurus they were even smaller. Mechanically they were not entirely useless: some estimates show that a T. rex could "curl" about 100 kg. However, that is only 1.25% of its body mass (something like a 200 lb human curling 2.5 lbs). So effectively they were useless in procuring prey.
Or at least in adults. Younger tyrannosaurids had proportionately longer arms while at the same time having proportionately weaker bite forces. As discussed below, these two may be ecologically linked.
How fast were tyrannosaurids? And how could we figure that out?
Some researchers have created computer models of Tyrannosaurus rex skeletons, applied estimates of muscle size and strength, and found that an adult could only run about 5.3 m/s (12 mph). But others using slightly different estimates find top speeds of 12-14 m/s (29-31 mph). For comparison, Usain Bolt's top speed was 12.4 m/s (27.7 mph). So adult Tyrannosaurus rex was either slower or faster than the fastest human so far... Part of the problem here is that we honestly don't know if we can accurate duplicate top speeds of living species directly from their skeletons, so at present these computer models should be treated with very large uncertainties.
Trackways can be used to reconstruct the speeds animals were traveling when they made them. A set of tracks of young Tyrannosaurus were found to record a walk of 2.25-3.47 m/s (5-7.76 mph). But any given tracksite is unlikely to record top speed, or even running speeds, since they sediment which records prints are by necessity going to be wet and sloppy: more solid surfaces are not in depositional settings.
An alternative to estimating absolute speeds is to evaluate relative degree of adaptation towards cursoriality ("running"). Within an ecosystem or within a clade we can compare which groups show more or less degree of cursorial specializations. Based on our observations of modern animals (and some basic mechanical theory), we recognize that cursorial species tend to have:
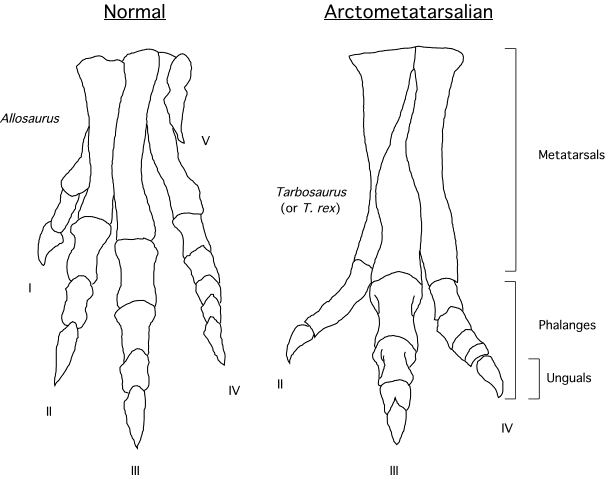
With the exception of shorter toes, tyrannosaurids show all of these compared to pre-tyrannosaurid tyrannosauroids, to other theropods (with a few important exceptions), and (especially) when compared to their potential prey. In particular, tyrannosaurids manifest a structure which I named an arctometatarsus (pinched metatarsus): proportionately slender metatarsals with the middle bone (metatarsal III) "pinched out" in the upper part between metatarsals II and IV, and forming a wedge-shape buttressed by the side bones below. In a new study, it was shown that tyrannosaurids additionally had specialized ligaments binding the feet in the buttressed region. The arctometatarsus was convergently evolved in the ostrich-like omnivorous ornithomimids (which similarly have disproportionately large hip muscles).
Notably, both tyrannosaurids and ornithomimids have the longest metatarsi at their body size:
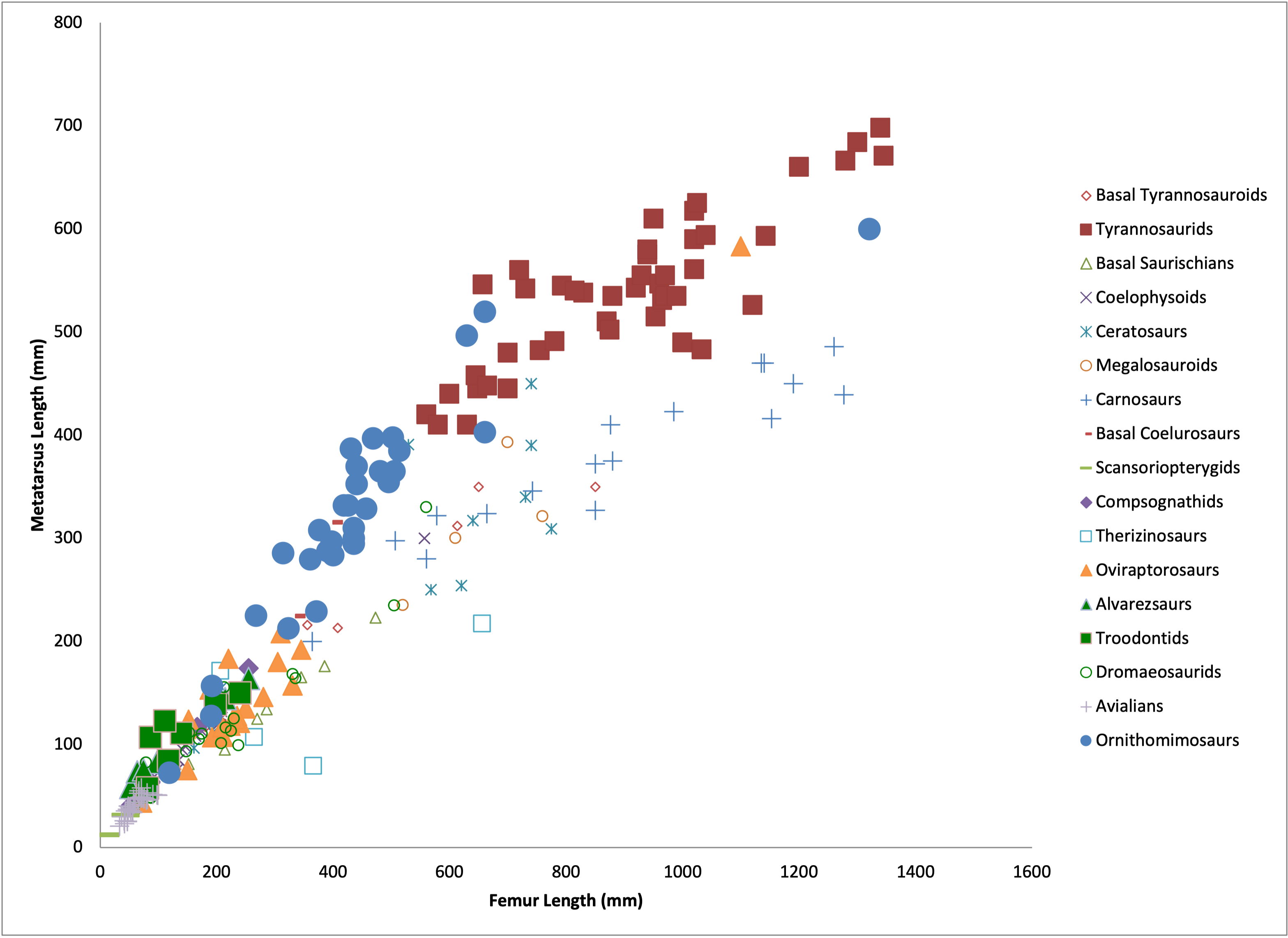
One thing that is clear from this is that young tyrannosaurids have the same limb proportions as ornithomimids (traditionally considered the fastest of Mesozoic dinosaurs.) And even giant tyrannosaurids have more cursorial proportions than other comparable-sized theropods.
More importantly for their ecology, however, is the fact that tyrannosaurids have much longer and more slender limbs prey of the same size. Tyrannosaurus and Albertosaurus didn't need to be fast as racehorses when their prey were relatively slow.
However, there is more to "cursoriality" than just maximum attained speed. Other issues include rate of acceleration; size of the home range; and even non-locomotory aspects such as the nature of the habitat or style of feeding. Another aspect of consideration is cost of locomotion. In a study of theropod locomotion we found that while longer legs resulted in clearly higher speeds in dinosaurs less than 1 tonne, in larger dinosaurs the longer legs didn't increase speed that much. However, it kept the energy required to move the body be lower. Thus, long legged smaller theropods were sprinters, and long legged large theropods were marathoners.
Another aspect of locomotion is agility (the ability to turn quickly). In yet another study we estimated agility by comparing the muscular power able to turn the body to the rotational inertia (the resistance of the body to turning). We estimated the latter by taking reconstructions of the body of theropods and dividing them into a series of slices, then estimated the density of each slice. The power for turning was determined by attachment area for muscles on the hips and tail. We found that tyrannosaurids had about twice the ability to turn compared than other large theropods of the same size. (And, as expected, we found that young individuals were much more agile than adults: this latter aspect plays out in the next section.)
Overall, the evidence points towards tyrannosaurids:
One common thread in tyrannosaurid research is that young individuals are dramatically different than the adults. We saw in the previous lecture, the huge size shift in dinosaurs during growth meant that they occupied different niches at each growth stage. Tyrannosaurids show a particularly strong case of this. Small individuals have slashing blade-like teeth, lower bite strength, proportionally smaller arms, greater agility, and much faster speeds; adults had crushing teeth, phenomenal bite strength, essentially unusual arms, relatively lower agility (although higher than those of other big predators), and relatively lower speeds. The data point to the younger animals were hunters of prey smaller than themselves, but as they grew up they shifted to attack prey which were about the same size as them.
A notable aspect of Asiamerica, though, is that it differs from earlier dinosaur communities in that there as essentially only one group of big theropods present (tyrannosaurids, of course). In the Jurassic and Early Cretaceous, there would normally be multiple different kinds of theropods present. For instance, in the Late Jurassic Morrison Formation of the American West, there were the giant megalosaurid Torvosaurus and allosaurid Allosaurus, the large ceratosaurid Ceratosaurus, the medium-sized stokesosaurid tyrannosauroid Stokesosaurus and megalosauroid Marshosaurus, and assorted taxa smaller than humans. In contrast, in Asiamerica there would be just one or two tyrannosaurids and a number of raptor dinosaurs smaller than humans. (In a few cases there are a few rare larger raptors.)
In the last study mentioned in this lecture, I compared the size distribution of predatory dinosaurs in these different communities to indeed confirm this size gap. I and others propose that young tyrannosaurids were effectively the mid-sized theropods of these environment, hunting prey that the adults could not catch in a fashion different than their adults.
Within their environments tyrannosaurids were more successful than other theropod groups had been earlier. But sadly for them, their tenure would be curtailed by a nasty rock from above.
A look at the issue of tyrannosaur ontogenetic niche shifts:

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}