


"In the animal world, there is the hunter and the hunted. Hounds and fox. Hawk and sparrow. Chicken and worm. We in the top most species have also joined the hunt. But who is to judge precisely which of us are the hounds or foxes as we enter... The Chase!." -- The opening text to the 1952-1953 American radio show The Chase
BIG QUESTION: What is the fossil record of animal defense?
Ecology: Energy in Living Systems
How does energy go from the non-living environment into the living world? One major way is photosynthesis. Here is the process greatly simplified:
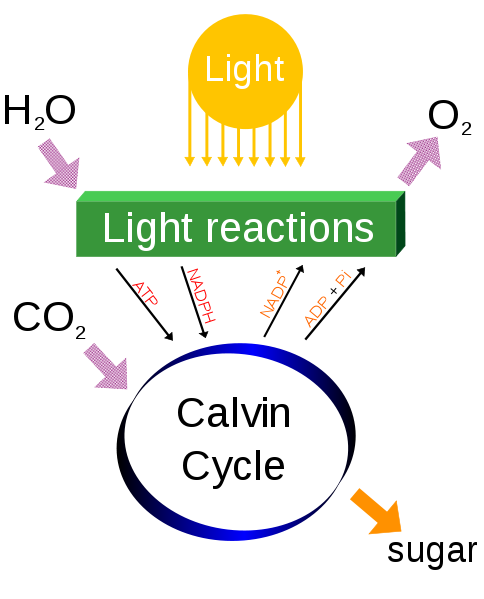
And here is the photosynthesis equation, simplified:
6CO2 + 6H2O + Sunlight yields C6H12O6 + 6O2
Or, in other words, photosynthesizers (plants and algae) take in water and carbon dioxide and using sunlight convert these into sugars and waste oxygen gas. Photosynthesizers are the major form of autotroph: organisms that don't eat other organisms.
In contrast, most other life forms (animals, fungi, most "protists", etc.) are heterotrophs: we get nutrients from digesting other life forms, primarily via the aerobic respiration equation:
C6H12O6 + 6O2 yields 6CO2 + 6H2O + Energy
(That is, food (glucose) plus oxygen yields waste carbon dioxide and waste water, plus energy).
In this reaction, we take in sugars and oxygen and release waste water and carbon dioxide.
So plants, algae, and other photosynthesizes are basically made up of air (and a little bit of other matter drawn from soil and/or water), while animals and other heterotrophs are made of food (ultimately plants) and air. This most important aspect of ecology is thus trophic relationships: who eats whom. These mark out relationships in a food chain of organisms that eat the organisms below them. However, since in nearly every case any species eats more than one type of food, trophic relationships are better displayed as a food web, comprised of many interlocking food chains.
The trophic level is the position an organism occupies of a food chain. Producers do not consume other animals: in terrestrial ecosystems, these are plants (which "eat" sunlight). Decomposers break down the bodies of any kind of organisms. Consumers are those that eat other organisms: 1st order consumers are herbivores (plant-eaters); 2nd order consumers eat 1st order consumers; 3rd order consumers eat 2nd and 1st order consumers; and so on, up to the apex predator (consumers that eat lower order consumers but are not typically eaten by any species but themselves).
As you move up a food chain, there is less and less energy available. Plants receive much (but by no means all) of the energy of incoming sunlight; some of that energy becomes stored as their tissues, but the rest of it is used for them to run their bodies. Herbivores consume plants, and gain some (but not all) of that stored energy, some of which they used to build their own tissues but the majority being used for their own operation (running, breathing, pumping blood, etc., etc., etc.). Predators eat the herbivores, gaining some (but not all) the stored energy in the tissues of the herbivores, using some to build their own bodies, but the rest for their operations. Thus, the amount of energy available at each trophic level becomes less and less: this is described as the energy pyramid.
One of the most important aspects of ecology (and paleoecology) is determining who ate whom: that is, reconstructing the food web. Sometimes there is direct trace fossil evidence: for example, this humerus of the Late Cretaceous Asian duckbilled dinosaur Saurolophus: the arrows point to bite marks generated by a tyrannosaurid (almost certainly Tarbosaurus, which is known from the same fauna).However, sometimes we do not have such direct evidence. In these cases, we can make relatively reasonable assessments. For example, it is unlikely that small theropods habitually killed the largest herbivores in their environment (while the latter were adults, at least: babies would be fair game), and it is unlikely that there were species that were not eaten by SOMETHING.
In the case of the herbivores, however, it becomes somewhat more difficult. For one thing, it is often the case that the conditions that are best to preserve plant fossils (especially of leaves and fruits) are not good for preserving bone, and vice versa. So we often lack good knowledge of the flora that co-existed with any fauna. That said, we have the digested plant remains in coprolites as positive evidence that dinosaurs ate particular plant species (although referring a coprolite to a particular dinosaur species can be difficult.)
However, as large-bodied terrestrial vertebrates, the above discussion is kind of biased. As we saw in the last several lectures, the ancestral state for Metazoa is NOT "kill your prey to get your food", even for second (and higher) order consumers. Most animals (ancestrally) are either suspension feeders or detritvores. In a sense, these are both examples of "scavenging", in that at least some of the food they are eating is already dead (and the eaters weren't the ones to kill it.) Instead, decaying bits of organisms (and fecal drops, and small living organisms) are swept up within the water column (for suspension feeders) or from the sediment (in detrivores.).
That said, there most assuredly ARE animals who make their living by actively killing their prey. And this predator-prey interaction is a major driver of evolutionary adaptations.
In some cases, organisms evolve in response to non-living aspects of their environment: different style appendages for moving over (or through) a surface; different types of integument to deal with various temperature conditions; etc. But very commonly organisms evolve in response to some aspect of another organism they are interacting with. In this situation, the other organisms themselves might evolve in response to the first set, and back and forth over the generations. This situation is co-evolution: two or more species evolve in response to new adaptations in the other.
Predator-prey interactions are a great example of co-evolution. Predators evolve new ways of finding, capturing, and killing their victims, and the prey evolve various defenses in response.
In this lecture we won't examine much in the way of predatory adaptions: the whole world of various jaws, claws, talons, fangs, venoms, snares, and so forth that meat-eaters have developed. We will instead focus on defenses. But we can start with one adaptation that works well for both predators and prey: eyes.
Predatory and Defense Adaptation: Visions: Eyes have evolved independently in many clades of animals. We vertebrates have eyes (in fact, eyeless vertebrates often seen weird), as do most arthropods and at least some mollusks (gastropods and cephalopods, most obviously). There are many other clades with eyes, but these are generally smaller and less well able to focus than in the examples listed above.
Eyes appear fairly early in the Phanerozoic record. For instance, Cambrian anomalocarids have been found with their huge, composite eyes (something like those of a dragonfly) preserved. Cambrian stem-vertebrates also show that eyes in the vertebrate lineage were already present. Examination of the early Cambrian Chengjiang Lagerstäaut;tte found that eyes were most common in predators and scavengers, and in nektonic or motile epifaunal forms.
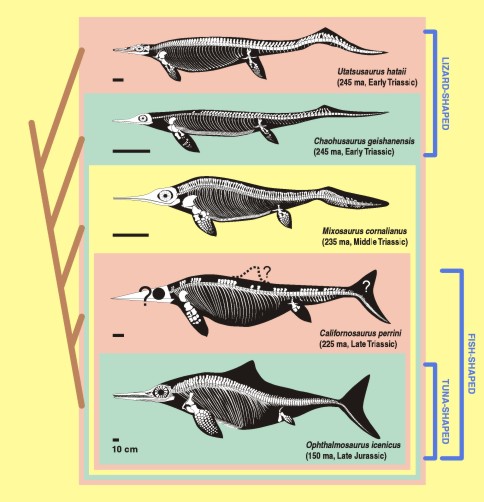
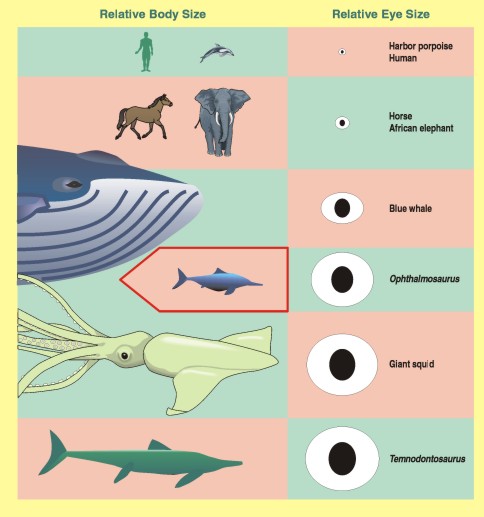
There are spectacularly large eyes within the Mesozoic marine reptile clade Ichthyosauria. Ichthyosaurs were among the most highly adapted of all reptiles to marine life (essentially the reptilian version of whales and dolphins.) But unlike cetaceans, their primary sensory mode was (like most reptiles) vision. Consequently to see in the darkness of the deep, they evolved the largest eyes known among animals. The larger the eyeball, the more photons it can catch: consequently, a bigger eye makes it easier to see at greater depths in the water. When you scale eyeball size against body size you find that the ichthyosaurs scale along the same trend as owls and king penguins (and giant squid), with the exception of the ichthyosaur Opthalmosaurus which had even BIGGER eyes for its body size than a typical ichthyosaur.
That said, good eyes are also useful for spotting approaching predators as well as looking for likely prey. We will see in a future lecture who they eyes of trilobites became very elaborate in the Paleozoic.
Defensive Adaptations: Armor: Such a ubiquitous adaptation that we have a whole evolutionary revolution dedicated to it, as we saw last lecture! (Well, to be fair, the Cambrian Explosion and the development of mineralized skeletons was more than just defensive armor, but that certainly is a major function.) The shells of typical shellfish (mollusks, brachiopods, hard-bodied crustaceans, etc.) serves a defensive function at least in part. Various vertebrate groups have developed body armor, include (but are not limited to):
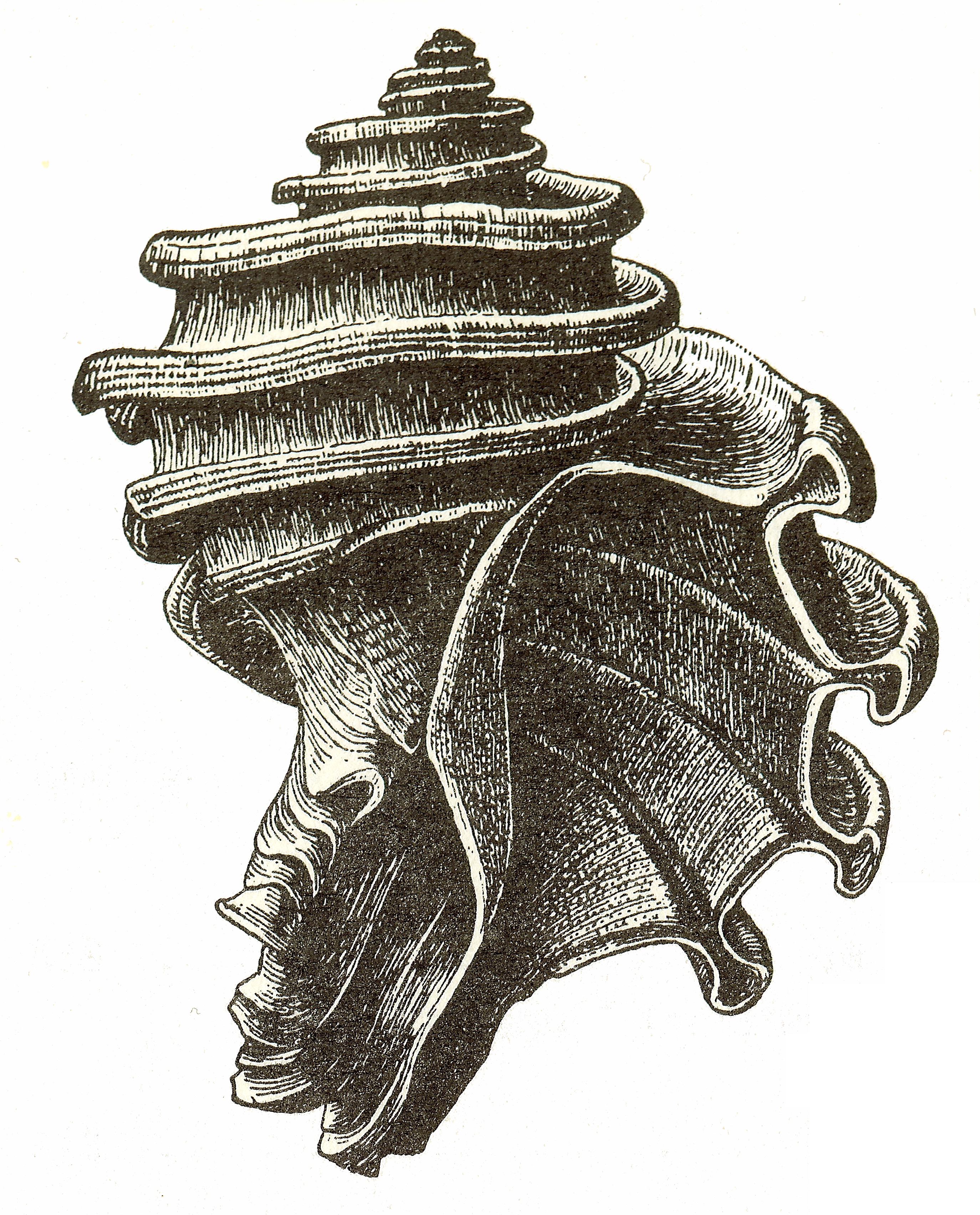
Response to Shells: Boring: The predatory response to armor is often to break it. Vertebrate predators might evolve crushing teeth to break through armor, for instance. But a particular strategy evolved by gastropods is boring. These snails drill into the shells (producing the marks seen on the fossils in the linked image) and either eat the shell out from the inside by sticking their proboscis in there, or by dabbing the victim with paralyzing poison, making it pop open.
Defensive Adaptation: Armor Enhancements and Weapons: A response to making your armor more difficult to break into is to use modified armor to keep the predator away. One way is to develop projections (spines, ridges, etc.) that make it harder for the attack to firmly grasp the shell: Maryland's state fossil, the Cenozoic gastropod Ecphora has such adaptations.
Alternatively, some of the armor might be transformed into weapons, like the spiked tail of stegosaurs and other tail weapons in mammals, dinosaurs, and turtles. Among non-armored weapons in prey species, cranial ornaments like horns and antlers are fairly common defensive structures. (It is worth noting, however, that observations of modern animals shows that weapons in prey items are often used as much or more against other members of the bearer's own species as they are against attacking predators.)
Defensive Adaptations: Camouflage: Sometimes the best way to be avoid becoming prey is simply not being protected. This might be simply hiding or staying still, or ducking down into a burrow or crevice. But natural selection has produced an even more elaborate version: camouflage. By evolving shapes, colors, patterns, etc. that match the surrounding, an animal can hope to avoid being seen.
There actually are examples of camouflage of this in the fossil record. Some examples include Cretaceous stick insects that mimic the leaves of the ginkgo trees they lived in or assassin bug, lacewing, and owlfly larvae gluing debris on their back to make them look unappetizing.
Defensive Adaptations: Escape: In the modern world, the masters of camouflage are the soft-bodied cephalopods (coleoids), such as octopods, cuttlefish, and squid. Not only can the change color and pattern almost instantly, but some can change the texture on the surface of their skin. As you might imagine, these soft tissue details are unlikely to survive, but a related trait is: ink.
When threatened, modern coleoids squirt out some dark colored ink; at the same time, they quickly change their body color to something lighter. This way the predator is likely to confuse the floating mass of ink as the coleoid itself. It turns out that ink sacs can actually be preserved in the fossil record: when found in extinct taxa, it strongly suggests that these extinct cephalopod clades were able to change their skin color. In contrast, no ink has been found in shelled cephalopods, like "nautiloids" and ammonoids: since the shell wouldn't change color--and since shelled cephalopods were generally not very fast--the distraction provided by the ink (even if they had it) wouldn't last long enough to help.
Of course, more common modes of escape are ducking down into burrows or crevices, or flight (in this context, flight as "getting the heck out of there", not flight as "flapping your wings" (although that can actually help)). The evolution of acceleration and speed adaptations in swimmers, terrestrial animals, and aerial animals is fairly well-studied. When we look at Cenozoic mammal evolution in a later lecture we'll examine the claims of co-evolution of predator and prey running adaptations.
Defensive Adaptations: Numbers: Yet another way that animals defend themselves is by living (or at least occasionally gathering) in vast numbers. Whether we call these schools, swarms, flocks, herds, or packs, the habit if being in a huge number of individuals means that the chance of any one individual will be attacked is much less likely. Gregariousness (the habit of living in groups) has evolved a great many times in the history of life.
The fossil evidence of gregarious tends to come from two main sources: body fossils and trace fossils. In the case of body fossils, monospecific assemblages (sites where a great number of fossils are found, and all of them are members of the same species.) These sites indicate that at the time of burial a great number of that species were present and died together, which itself strongly suggests that (at least on occasion) they lived together in such groups in live. And as with most cases in the fossil record, more instances of the same type of site suggests that this was indeed a regular feature of the biology of that species, and not just some unusual freak occurrence.
An alternative line of evidence is monospecific track sites. These would be outcrops with a great number of traces of the same species. Since trace fossils are produced by living organisms, they can't be transported to the same spot postmortem (which is a potential issue with at least some body fossils.) Granted, it is difficult to determine whether all the tracks formed at the same time, or whether they accumulated over time. (This is less of a problem for terrestrial deposits: the sediment would only be wet for a short period of time, so all the tracks would have only this interval in which to be formed.) There are many cases (from marine invertebrates to terrestrial mammals and dinosaurs) of monospecific tracks.
Another type of trace fossil for group behavior is nesting colonies. Nesting colonies are a special case of gregariousness, where many animals lay their eggs in a relatively small area. In such colonies it is more difficult for a nest thief or other predator to sneak in and attack and egg or baby without being spotted. Nest colonies have been found for a number of fossil animals, including several different groups of dinosaurs, pterosaurs, and a few others.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}