


I come from the water
I crawled upon the shore
I left my brothers there
I got what I came for"
-- Todd Lewis, lyrics for "I Come From the Water" (1994) by The Toadies
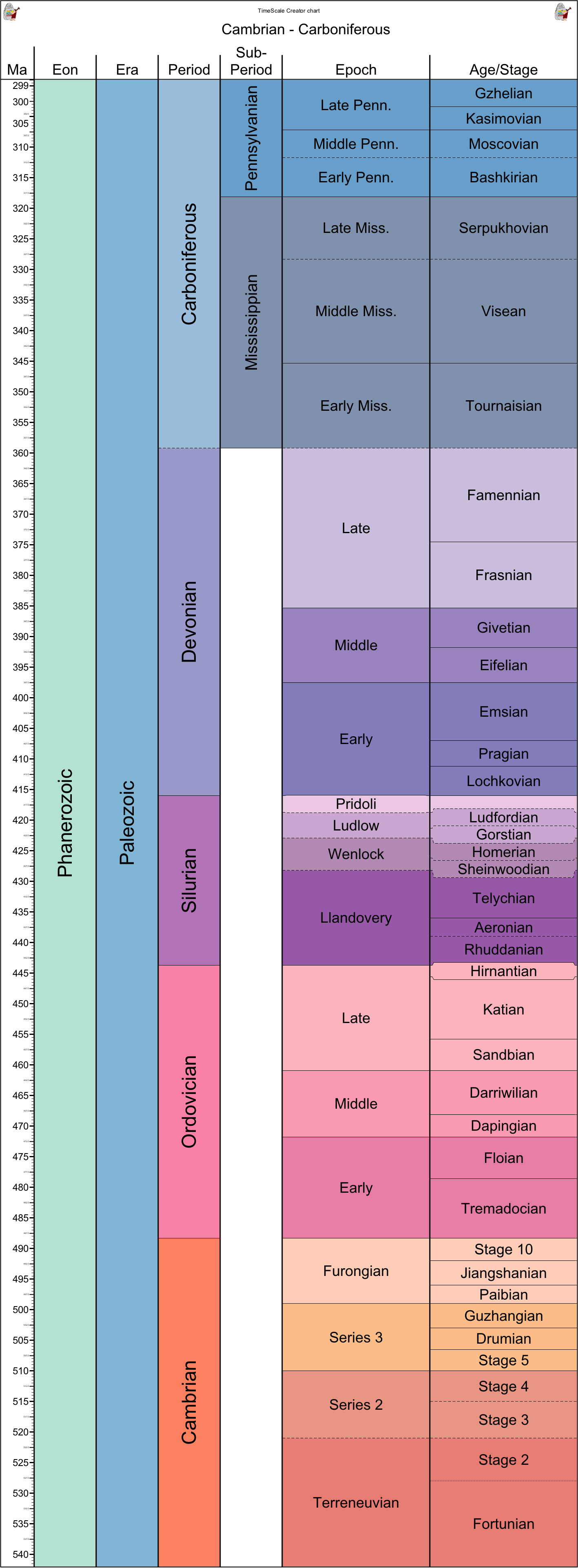
BIG QUESTION: What were the stages in the rise of life on land?
A Land Planet
The Earth may be covered by 70% water (almost all of it ocean), but most of the life is not in the seas. A recent (2018) study estimated there was 470 gigatons (Gt) of carbon made up of land organisms (450 Gt of that is plants), compared to only 6 Gt of marine life. (By the way, there is about 70 Gt in deep subsurface bacteria and archaeans). When only animals are considered, the marine realm does dominate, but not to the same overwhelming degree as the overall pattern (for those interested, 2.1 Gt in marine animals versus 0.589 Gt in terrestrial).
In terms of species diversity the terrestrial realm also greatly exceeds that of the marine realm: it is estimated that between 85-95% of all living macroscopic species live on land.
Yet this wasn't always the case. Life arose in the seas, and for the majority of Earth's history it was limited to the marine realm. Only a few clades have adapted to life on land.
When and why did this change take place? That is the subject of the present lecture. After reviewing some basic principles, we focus on three of the most successful groups to colonize the land: land plants (embryophytes), arthropods, and land vertebrates (tetrapods).
Timescale for the lecture:
Why would organisms ever abandon life under the sea? After all, both freshwater and the terrestrial realm (together the continental environments) are extremely challenging to organisms who first evolved in salty water.
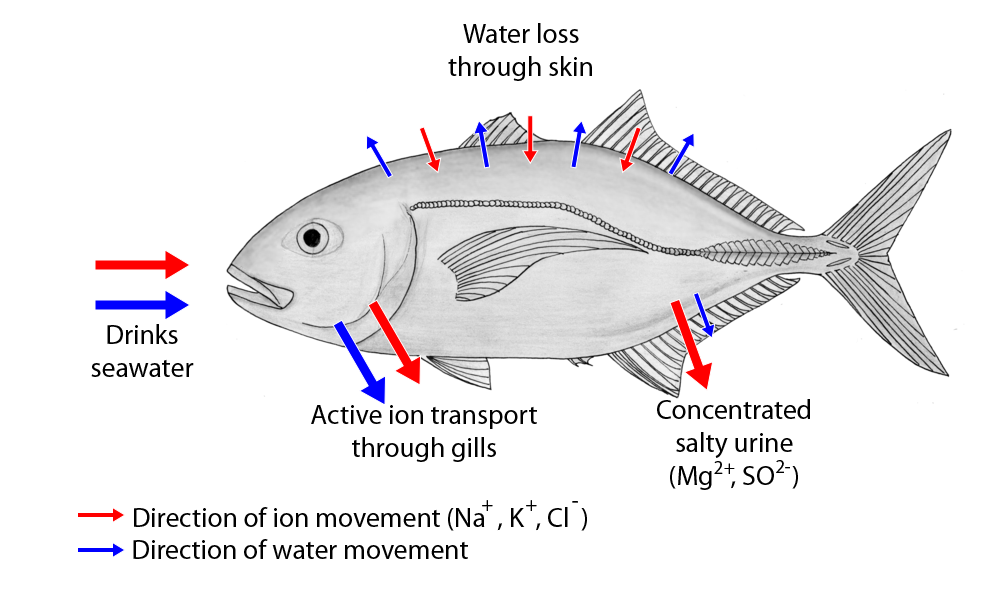
Not every organism can tolerate different salinity conditions. Osmoconformers have very little "buffering" between internal physiology and the external environment. Internal salt levels therefore tend to match the ambient conditions. Since organismal physiology requires salts, it is impossible for osmoconformers to survive well in freshwater conditions, because they cannot maintain their internal osmolarity.
In contrast, osmoregulators have mechanisms (normally associated with the excretory system) to control salt levels. Freshwater osmoregulators have higher internal osmolarities than ambient conditions, and so need to excrete excess water to retain their salt levels. In contrast, saltwater osmoregulators, the internal osmolarities are lower than ambient, so they need to retain water but excrete excess salt. Given these different requirements, most animals are stenohaline (have narrow range of salinity tolerances). However, some are euryhaline (have broad range of salinity tolerances). Not surprisingly most groups that make it to freshwater and land tended to be euryhaline.
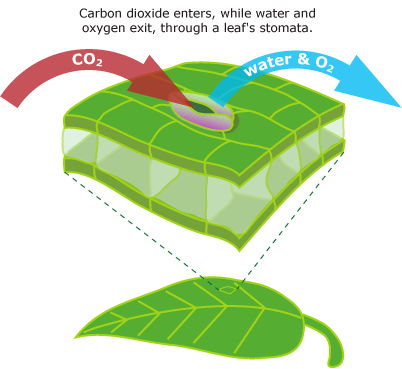
Life in freshwater can be fairly difficult:
Even tougher is life out of water, where organisms have to deal with
There are certain similarity in the traits required to live in both freshwater and terrestrial habitats compared to marine life:
Estuaries (bays, deltas, wetlands, etc.) seem to be ideal settings for the transition from the marine realm to continental ones:
The trace fossil record of estuaries show from the Ediacaran through the Ordovician ichnofossils of marine organisms (especially of arthropods) in low energy, fine-grained bay deposits. During the Silurian, Devonian, and Carboniferous there is an increase of non-arthropods (bivalves, polychaete annelid worms, etc.) and presence of fossils in all estuarine environments.
Some major groups of organisms (red algae, corals, brachiopods, echinoderms) never made it into freshwater. Many of these are calcifying osmoconformers, and thus cannot handle the particular osmolarities of freshwater. Others made it into freshwater but not onto land: non-coral cnidarians (like hydroids), bivalves, sponges, bryozoans. Nearly all of these are suspension feeders, and thus they could not feed in the air.
But there are many groups that have made it onto land: earthworms from aquatic annelids; pulmonate and other land snails from aquatic gastropods; and the Big Three that will be the focus of this lecture:
For these, let's examine:
Even before the colonization of the Big Three, there were early phases of colonization (although limited) on land:
Land plants are the sister taxon to the freshwater green algae Charales (stoneworts), and together are nested among freshwater green algae. Charales have "stems" which are multinucleate single cells, not homologous to the vascular stems of tracheophytes (see below). Charales reproduce by sending flagellated sperm cells in the water, and indeed spend their entire life cycle in the water.
In contrast the most primitive living embryophytes ("bryophyte-grade" forms) reproduce by releasing spores into the air, which land on wet surfaces, producing a generation of tiny plants that makes sex cells. The sperm are carried along by raindrops or over wet surfaces, fertilizing the egg cells, which grow into an adult plant which can generate spores.
The oldest possible embryophyte remains are the so-called "cryptospores" of the Cambrian and Ordovician: these are spores which are identical in form to basal crown-embryophyte spores, but could conceivably be from some aquatic stem-embryophyte taxa (i.e., that lived in the water but spread spores through the air to colonize new water habitats). There are also some possible thalloid (multicellular, tissue-grade but no definite organ systems) algal from the Cambrian and Ordovician which are conceivably from some terrestrial basal plants. Also present in the Ordovician, however, are tissue of organ-grade embryophytes.
They basal modern land plants (liverworts, hornworts, and true mosses: see cladogram above) are all built very low to the ground, lacking significant support or the means to transport fluids up into tall tissues. However, they do contain two major adaptations required by all land plants:
The next major phase of embryophyte evolution is Tracheophyta, the vascular plants. These plants show new adaptations:
The oldest tracheophytes are the paraphyletic grade of "rhiniophytes", known first from the Early Silurian. These primitive land plants show stems, branches, sporangia (spore-generating organs), but no leaves.
The next major adaptation of plants is:
The primitive Silurian and Early Devonian leafed plant Baragwanathia is a member of the Lycophyta (clubmosses). Lycophytes (in the form of lycopsid "scale trees") were one of the first very successful land plants, eventually producing trees up to 30 m tall. However, they still reproduced by spores and thus needed to live in environments that were at least seasonally wet.
The leaves of lycophytes ("microphylls") are very small and simple. The next major adaptation in the plant conquest of land were large complex vascular leaves. One of the first branches of the Euphyllophyta ("good leaf plants") are the Monilophyta (ferns and horsetails). Ferns still reproduce by spores, in a manner similar to "bryophyte-grade" plants. Ferns are actually quite diverse, producing "classic" ferns, tree ferns (in which many stems growing next to each other support the tall structure), and horsetails (a major group of ground cover plants in wet environments, and during the late Paleozoic a major group of tree-sized plants).
The last major advance in the first wave of plant colonization of land is the development of wood (tough tissue with bark and the ability to grow in thickness as the plant grows in height). The oldest members of Lignophyta (woody plants) still reproduced with spores: this differs from all living woody plants which are also members of Spermatophyta (seed plants). Early lignophytes are present in the Devonian, and were the trees that dominated the Devonian forests.
Today there are many diverse groups of panarthropods that come onto land:
Unlike embryophytes (a clade) or the tetrapods (a clade), land arthropods are not a monophyletic group. Instead, they came onto land in numerous independent waves. This is because the arthropod body plan had numerous features easily adapted to life on land:
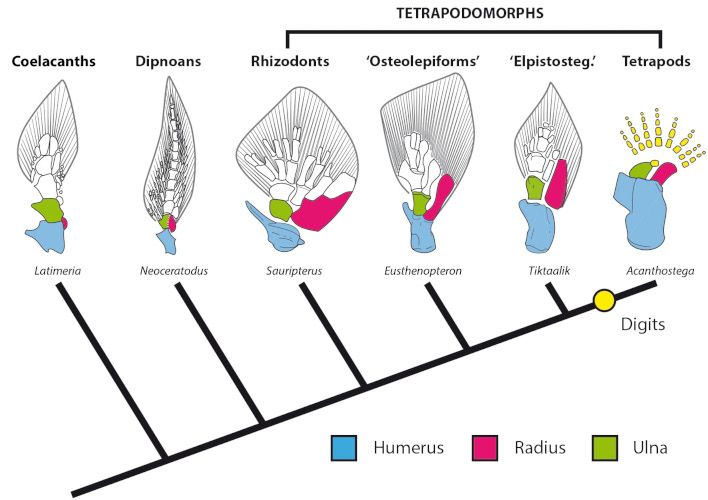
The Devonian is often called the Age of Fish because of the great diversity of different lineages present. Among these were the sarcopterygians (lobefins), which had inherited from their ancestors:
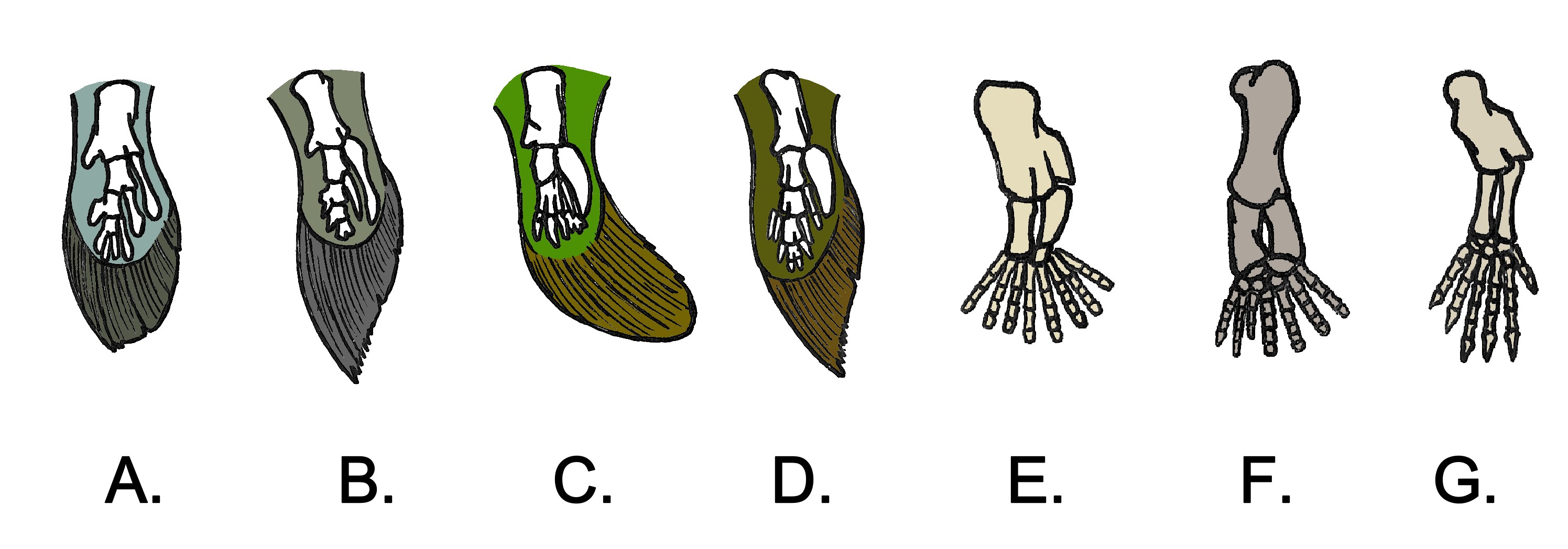
Furthermore, sarcopterygians themselves had fins with bones running down the body of the fin, making it very strong and muscular. This can be used to dig a hole (as some lungfish use in order to survive the dry season); to push through vegetation; and to crawl out of one pond and move into another. (This latter ability isn't that strange: it is found in some living actinopterygians such as catfish, mudskippers, snakeheads, even freshwater eels).
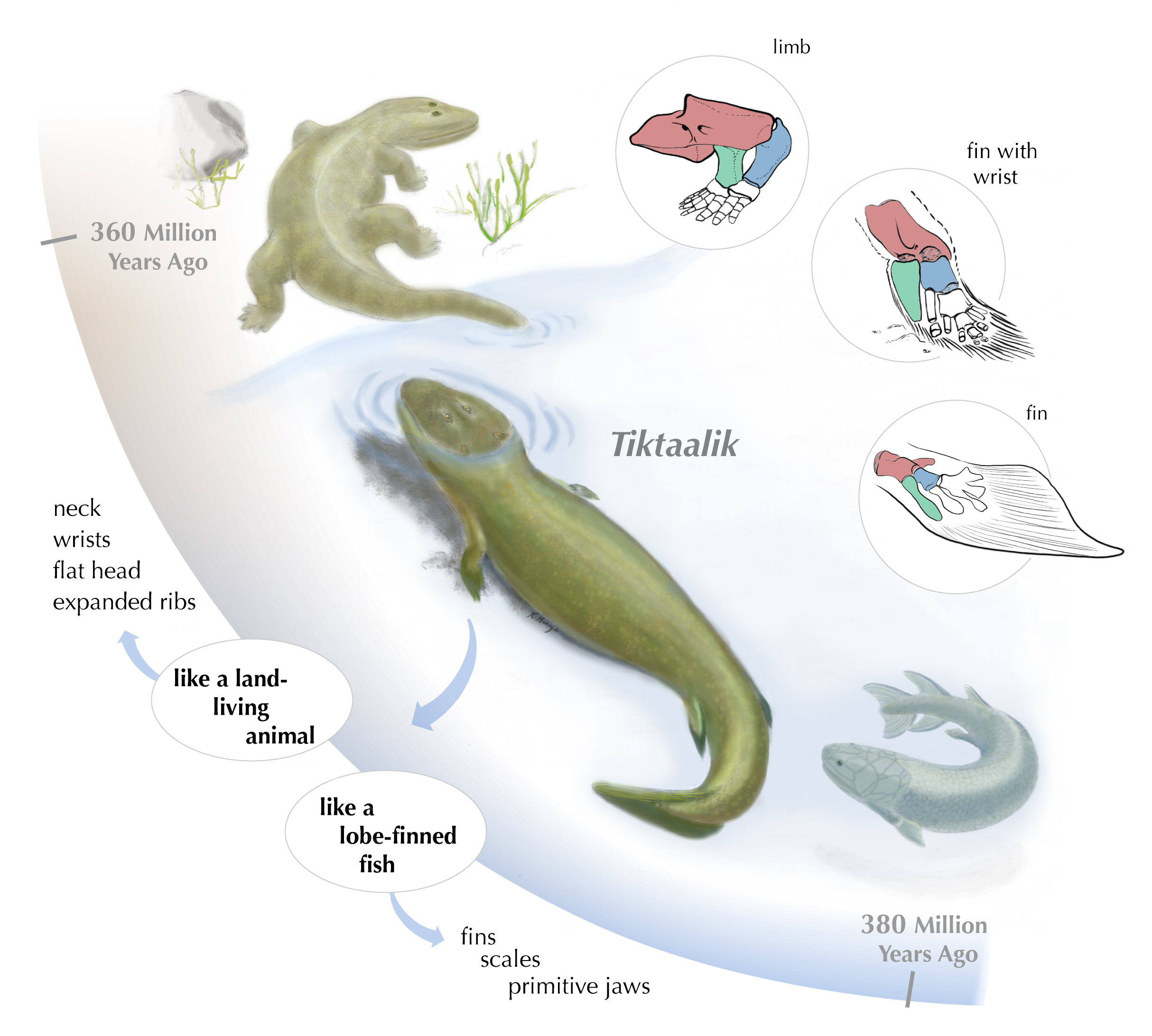
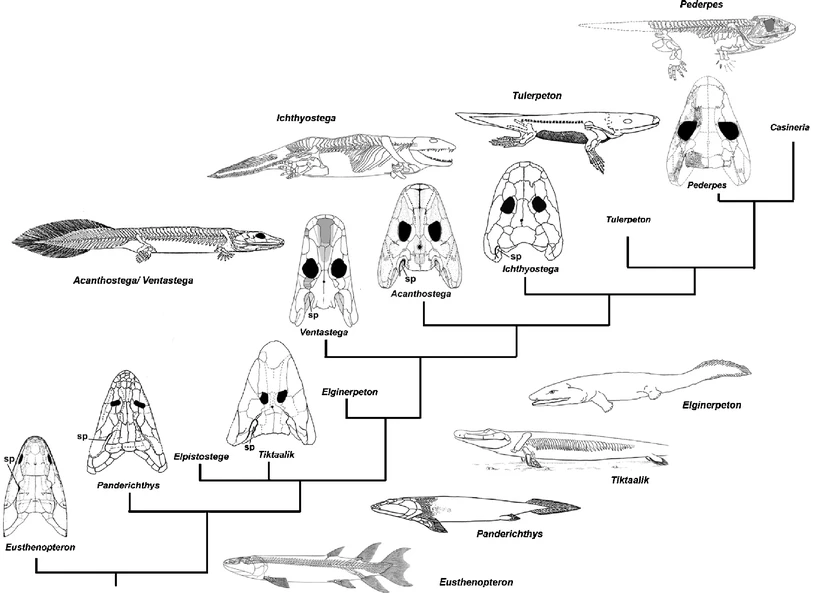
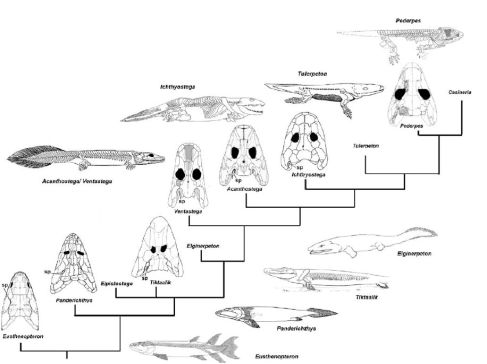
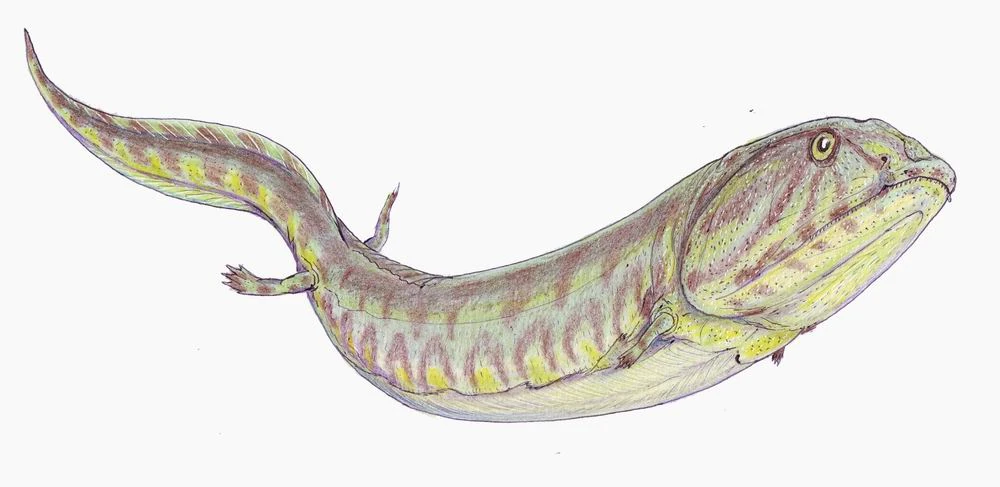
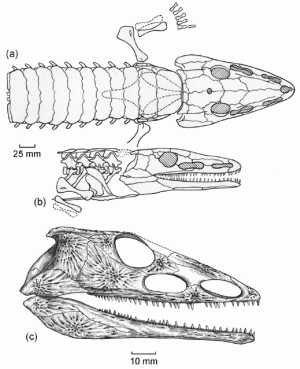
All the major groups of sarcoptergyians (Tetrapodomorpha) is of particular interest for us. Like the other groups (coelacanths and lungfish), some were marine and some were terrestrial. In fact some of the aquatic tetrapodomorphs were large shark-like predators. Among them were some in which the lobe fin developed wrists and elbows and in which the skull became flatter and wider. These suggest these advanced tetrapodomorphs spent more time in shallow water, even shuffling out of the water in order to catch food on the land. The best known of these "fishapods" (Tiktaalik of the Middle Devonian, shows that it still lacked fingers and toes (digits), but otherwise was very similar to the next phase of vertebrate evolution.)
Stegocephalia is the clade of fingered vertebrates. In these forms the lobefin fin lobe became the walking limbs. However, basal stegocephalians like Acanthostega and Ichthyostega were still primarily aquatic.
In fact, they were built primarily for swimming, and could probably only move around on land as well as a modern mudskipper or snakehead fish:
Traditionally the hypothesis was that stegocephalians developed better limbs in the context of freshwater (since ponds are more likely to dry up than the ocean). While this is still pretty reasonable, the oldest stegocephalian tracks (from the earliest Middle Devonian) are on a marine beach, not a lake shore. So it is possible that terrestrialization was straight from the sea, not from freshwater.
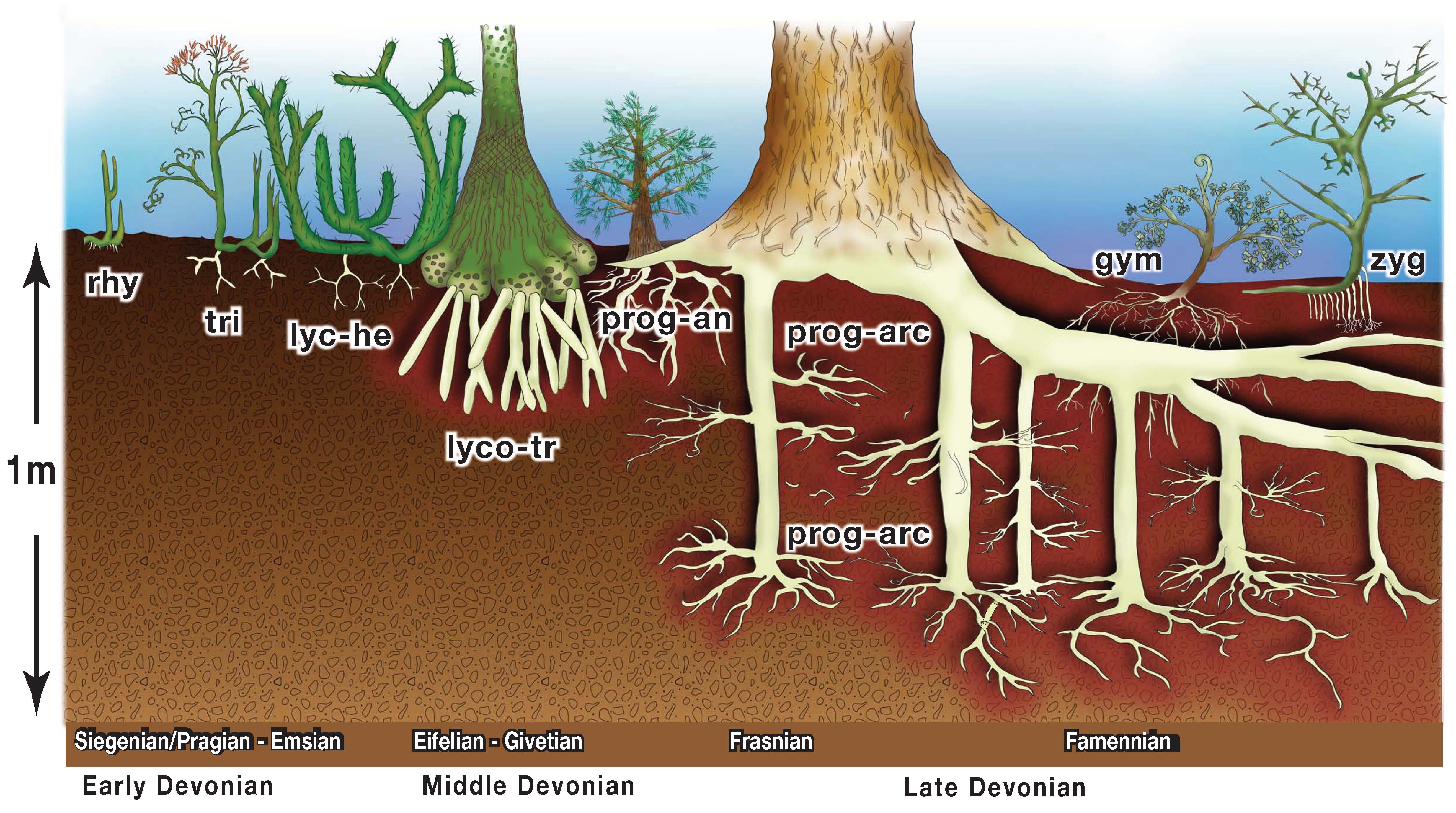
During the colonization of land, it wasn't just animals and plants adapting to land condition. Biological activity actually modified the surface (and subsurface):
During the Late Devonian, upland communities developed, with tracheophytes as the producers, arthropods as the major herbivores and predators, and fungi and arthropods as the detritivores. Stegocephalians were not a major part of this community, and even the plants couldn't move too far from the water source until the rise of seeds.
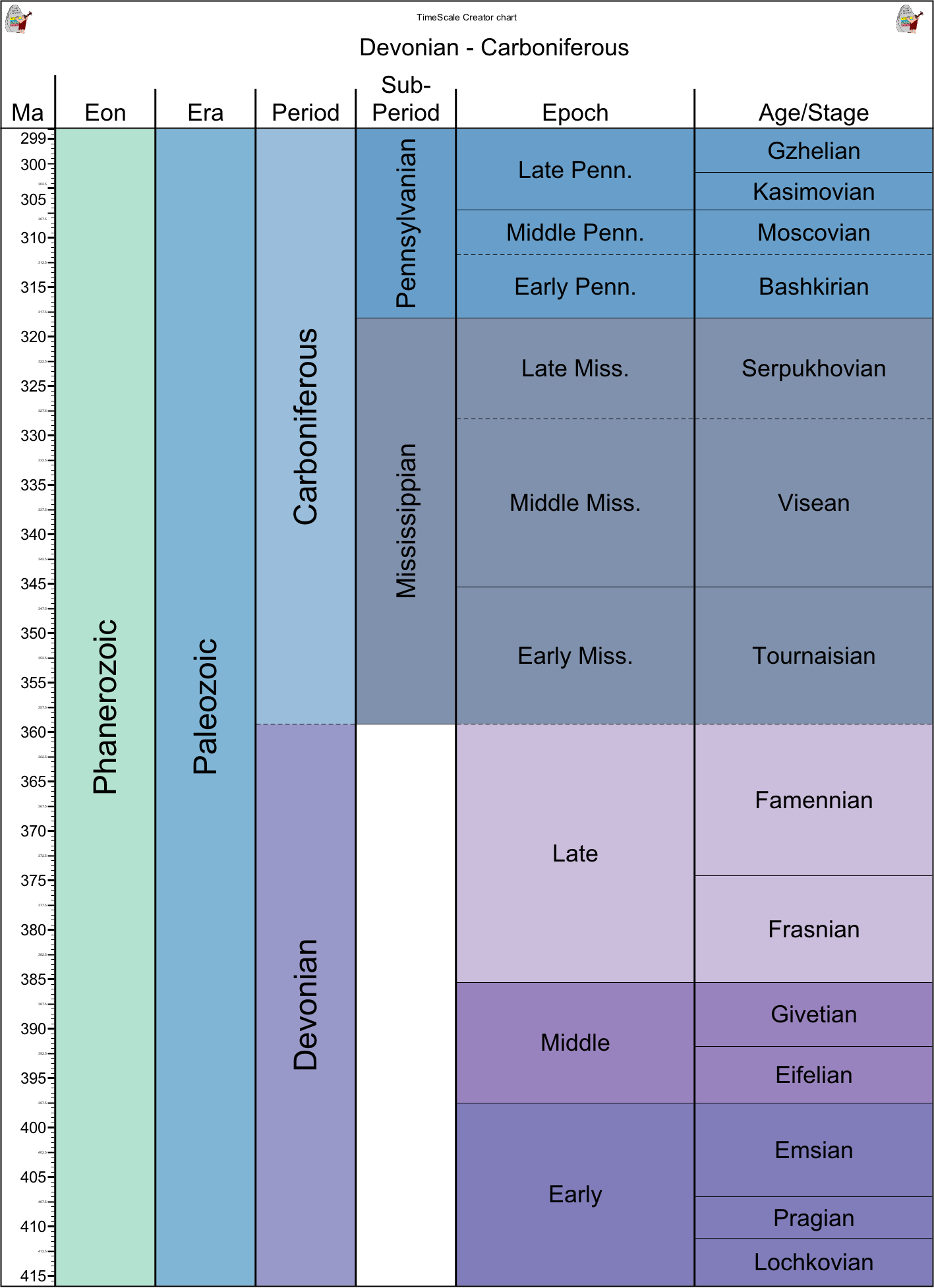
Romer's Gap: During the mid-20th Century, vertebrate paleontologist Alfred Sherwood Romer noticed that although basal stegocephalians show up in the Late Devonian, but it wasn't until the Pennsylvanian Subperiod that the more advanced Tetrapoda appeared. Over the last couple of decades a few stegocephalians (including early tetrapods) have been found in the Mississippian Subperiod, but not terribly many. Later work suggested that this pattern appears in arthropods as well, with early appearance in the Devonian but major diversifications in the Pennsylvanian.
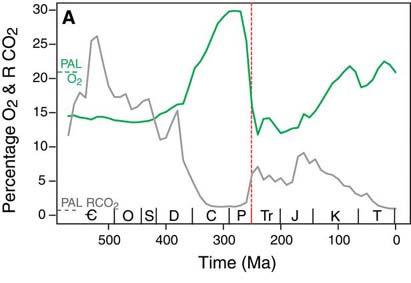
Is Romer's Gap real, or is it an artifact of poor sampling during the Mississippian. While it is true that some additional fossils of stegocephalians and arthropods have been found in this Subperiod, the discrepancy between initial appearance and much later diversification seems to hold. Some have suggested low oxygen levels in the atmosphere may have suppressed diversification, and that only with the rise of increased oxygen did the groups increase in specialization.
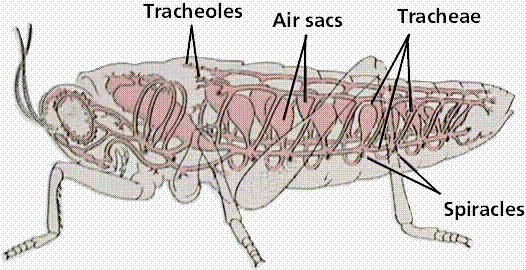
As possible evidence for the decreased oxygen level during the Mississippian, the size of the spiracle (first gill slit in jawed vertebrates, modified into an opening in the back of the skull used to help flush oxygenated water past the gills) increased in sarcopterygians. Since the oxygen level in the water is controlled by the atmosphere, this may indicate the need to increase the amount of water needed to keep the body oxygenated.
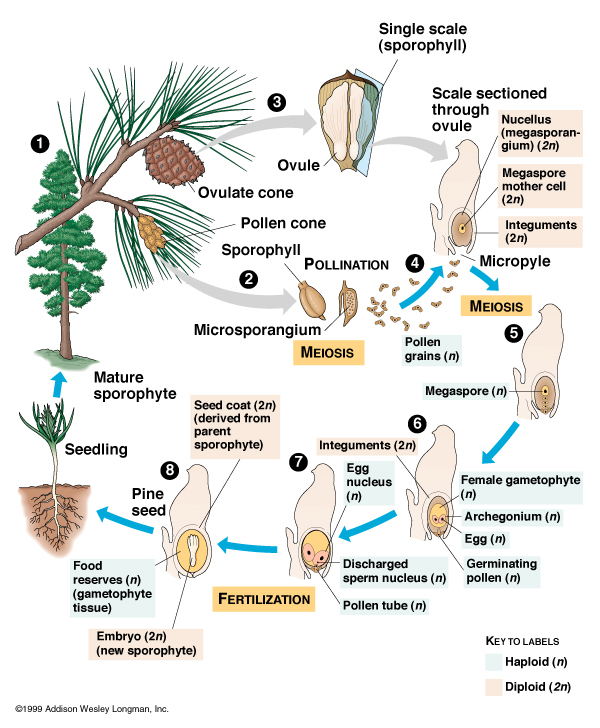
Seeds appeared among lignophytes during the Devonian. The seed plants (Spermophyta) today include the non-flowering Acrogymnospermae and the flowering Angiospermae. The term "gymnosperms" refers to the paraphyletic grade of non-angiosperm spermatophytes: this includes the acrogymnosperms, the non-flowering stem-angiosperms, and a set of basal clades collectively called "pteridosperms" ("seed ferns", because of their leaf shape).
In spermatophytes, the old spore plant life cycle is replaced with a new system. In the gymnosperm condition, the adult plant produces male cones (that produce pollen, spread primarily by wind), which will land in the female cones, which fertilized the egg, to produce a seed. The seed has a cover which allows it be deposited on much drier land than any spore plant could colonize. Thus, seed plants were able to colonize essentially all habitats on land except for the driest deserts and coldest polar regions.
Late Carboniferous Forests: the forests that developed by the Late Carboniferous (or Pennsylvanian Subperiod) are very well-studied, as these are the major environment in which good quality coal was generated (more about this below). The lowlands were dominated by lycopsid "scale trees"; slightly upland from those were calamite horsetail trees; further up from that tree ferns and seed ferns; and seed ferns and other primitive seed plants dominating the land farthest from water.
Completing the Conquest: Wings
During the Carboniferous arthropods continued to diversify, including new varieties of arachnids and the arthropleurids (or "godzillapedes"), the largest land-dwelling arthropods of all time. But the primary new adaptation of this part of arthropod history is the origin of insect wings. Winged insects--Pterygota--may have actually be present in the Devonian (if Rhyniognatha is in fact a pterygote); however, there are definite winged insects in the mid-Carboniferous (just above "Romer's Gap").
Debate remains whether wings were derived from lateral extensions of the thoracic body surface (paranotal surface) or from the breathing-part of the limbs (exites). Wings may have initially helped with cooling the body and/or sailing across water surfaces, but of course their main function is to turn the world of the insects into a three-dimensional world.
The most primitive wings were fixed-wings: not fixed like those of an airplane, but ones that at rest are either stuck out to the sides or held largely vertically over the body (as in modern dragonflies in the first case, and damselflies and mayflies in the next). They could do lots of flying and maneuvering around, but fixed-wing insects are limited in moving through dense vegetation, burrowing, and so forth.
Folded wings are characteristic of the Neoptera (the most diverse group of animals of all time). Folding wings allowed insects to colonize many more habitats than basal pterygotes.
An additional set of changes associated with insect evolution is the transformations during ontogeny. Basal pterygotes have nymphs that are wingless and don't look very much like the adults. It is tempting to think that because most dragonflies, damselflies, and mayflies have aquatic nymphs and terrestrial flying adults that the ancestral flying insect condition was an amphibious form. But the breathing structures of the different nymphs are non-homologous, and the even more basal non-flying insects and non-insect hexapod (and even some primitive dragonflies) is to lay eggs in moist soil rather than water.
Neopterans almost all have terrestrial (or burrowing) young. Basal neopterans have nymphs that look like non-flying adults (no major metamorphosis). Derived neopterans (Endopterygota) have are holometabolous (have complete metamorphosis): their larvae (maggots, caterpillars, grubs, etc.) look nothing like (and have ecologies nothing like) the adults.
Completing the Conquest: Tetrapods, Including Amniotes
During "Romer's Gap" there are a few new branches of stegocephalian, including 2 m or longer eel-like aquatic Crassigyrinus. But the most successful group of tetrapods to appear during this time are the Tetrapoda (the crown-group of Tetrapodomorpha). Tetrapods are distinguished from other tetrapodomorphs by lacking gills in the adult, having a five-fingered hand, and a distinct neck (complete separation of the skull from the shoulder blade.
Todays living tetrapods are the Lissamphibia (frogs, salamanders, caecilians) and the Amniota (mammals and sauropsids). In the Paleozoic, however, there were many other branches; some (Batrachomorpha, the Lissamphibia total-group) closer to the living amphibians, and the others (Reptilomorpha, the Amniota total-group) closer to the amniotes. All of the batrachomorphs (and the Carboniferous stegocephalians outside Tetrapoda) and some of the reptilomorphs were "amphibians" in the broad sense: that is, they had aquatic "tadpole" larvae with external gills (non-homologous to the internal gills of "fish"), but terrestrial adults.
There was a tremendous diversity of Paleozoic "amphibians": alligator-like forms, boomerang-heads, snake-like forms, armor-plated forms, and more. There were even scaled, marine amphibians (totally distinct from the naked-skinned, salt-intolerant lissamphibians).
All these forms, however terrestrialized as adults, were still stuck living near water in order to reproduce. Tetrapods were capable of the final stages of terrestrialization through the amniotic egg. These eggs have:
Amniotic eggs thus freed amniotes from the pond, and allowed them to colonize all terrestrial habitats. In this way they are exactly analogous to the seeds of spermatophytes.
There were additional traits that unite the amniote groups, such as:
For all their diversity, there is no evidence that any of the other groups of tetrapodomorphs included herbivores. In contrast, many amniote groups (and the immediate outgroups to crown-group Amniota, which may have had amniotic eggs) evolved herbivory in the Permian. From this point on, amniotes joined arthropods as major feeders on land plants.
One of the main changes in anatomy of the amniotes (and other tetrapod) was the development of hearing adapted to the air: impedance matching ears, in which a large tympanum ("eardrum") collects sounds that get delivered to inner ear by one or more bones of the middle ear. Some modern tetrapods lack an eardrum, like salamanders, and the fossilized skulls of basal amniotes and indeed most basal tetrapods show that they lacked these structures. But frogs, mammals, turtles, and eureptiles (lepidosaurs, archosaurs, and their extinct kin). Based on fossil evidence, these groups (and an extinct group of parareptiles) evolved this trait only in the latest Pennsylvanian or early Permian
The Carboniferous ("coal bearing") gets its name from the fact it is the period when most of the coal of the planet formed. Coal formed in low-lying well-watered forests (coal swamps) in which plant material was buried before it could be decayed. Coal seams stretch for hundreds of kilometers. They are found in repeated cycles of non-coal coastal sediments and coal seams, called cyclothems.
Why the repeated cycles? The initial development of soils and early coal swamps sequestered a considerable amount of carbon in sediment, lowering atmosphere CO2 and global temperature. In fact, it became so low (comparable to modern values, and much lower than most of Earth history before or after) that the Earth entered an "icehouse condition", and even further into an "ice age" condition. The Milankovitch cycles that dominated later ice ages were very much in effect in the Carboniferous: indeed, this is why there are cyclothems, as the rise and fall of glaciers produced repeated burial of swamps. Carbon kept getting sequestered, and as a consequence carbon dioxide levels were kept very low.
A consequence of the burial of unoxidized carbon and the massive primary productivity of the coal swamps was an increase in the level of atmosphere oxygen (the highest it has ever been). This increased oxygen level probably explains the existence of terrestrial arthropods of unusual size (like arthropleurid "godzillapedes", giant pterygote insects, and large terrestrial eurypterids. One of the main limitations on modern arthropod size is the fact that arthropods do not have powered respiration, but rather rely on passive diffusion of air over their book lungs or into their spiracles and through their trachea. With the increased levels of oxygen, much larger arthropods could be supported. With reduced oxygen levels in the later Permian (as well as the increase in diversity of amniotes), the arthropods were kept at smaller body size.
Colonization of land produced substantial gains in Life's ability to access nutrients from the Earth System. In modern ecosystems, observations show that total primary productivity per unit area is about 4 times greater on land than at sea. (Because of variation on which organisms are involved and the variations in temperature and atmosphere, we cannot say yet whether this ratio has remained comparable throughout the post-Silurian world).
In the Permian Period (the final period of the Paleozoic), plants and arthropods were as fully terrestrial as they would ever become, but the final terrestrialization of the tetrapod community occurred. While the bodies of Carboniferous amniotes were essentially fully terrestrial, their ecosystems were not. As 20th Century paleoecologist E.C. Olson noted, Late Carboniferous and Early Permian tetrapod communities still received most of their energy from the water: algae feeding fish, which fed aquatic amphibian-grade tetrapods, which fed amniotic predators. There were a few terrestrial herbivores, including some larger ones, but these were numerically much rarer than the terrestrial predators: this seeming contradiction to ordinary ecological rules stems from the fact that most of the energy among the larger animals was being derived from aquatic ecosystems.
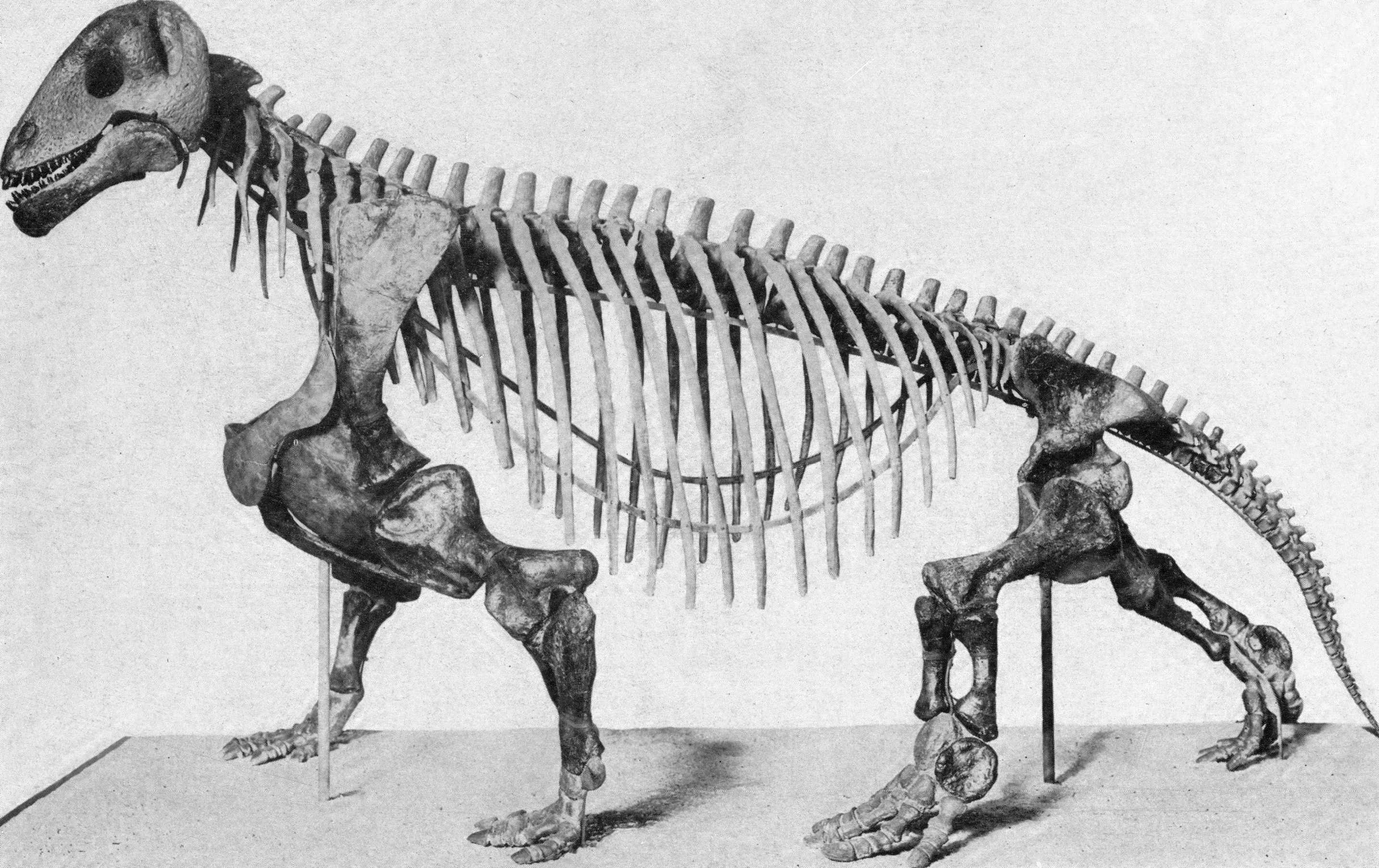
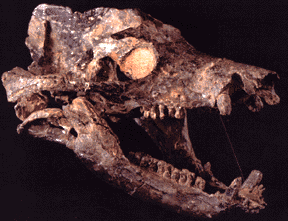
The Middle and Late Permian saw a change in this. The more derived members of the synapsid lineage of amniotes arose: the Therapsida. Once called the "mammal-like reptiles", they are not true reptiles. Instead, they are the advanced branch of the synapsid phylogeny. They differed from earlier synapsids by having teeth divided into nipping incisors, biting canines, and grinding cheek teeth (an arrangement that should sound familiar, as it is what is in your mouth!) and Forelimbs more powefully developed than hindlimbs. The Middle and Late Permian therapsids included Dinocephalia (large slow-moving herbivores and carnivores of the Middle Permian; Gorgonopsia, the apex predators of the Late Permian; Dicynodontia, the extraordinarily successful beaked, two-tusked burrowing omnivores and herbivores (which would persist well into the Triassic); and the oldest Cynodontia, initially smaller specialized carnivores and omnivores which eventually produced the mammals.
These therapsid protomammals, and their Middle and Late Permian sauropsid contemporaries, evolved ecosystems much more familiar to those of the Mesozoic and Cenozoic. Large predators were rare, and medium-to-large herbivores were common. Diverse modes of herbivory evolved among these animals, as embryophyte vegetation increasingly became the base of the terrestrial food pyramid.
Such was the terrestrial ecosystems at the end of the Paleozoic Era. These Permian animals were unsuspecting that beneath their feet, danger grew to destroy the world they knew...

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
_male_3.jpg){kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}