
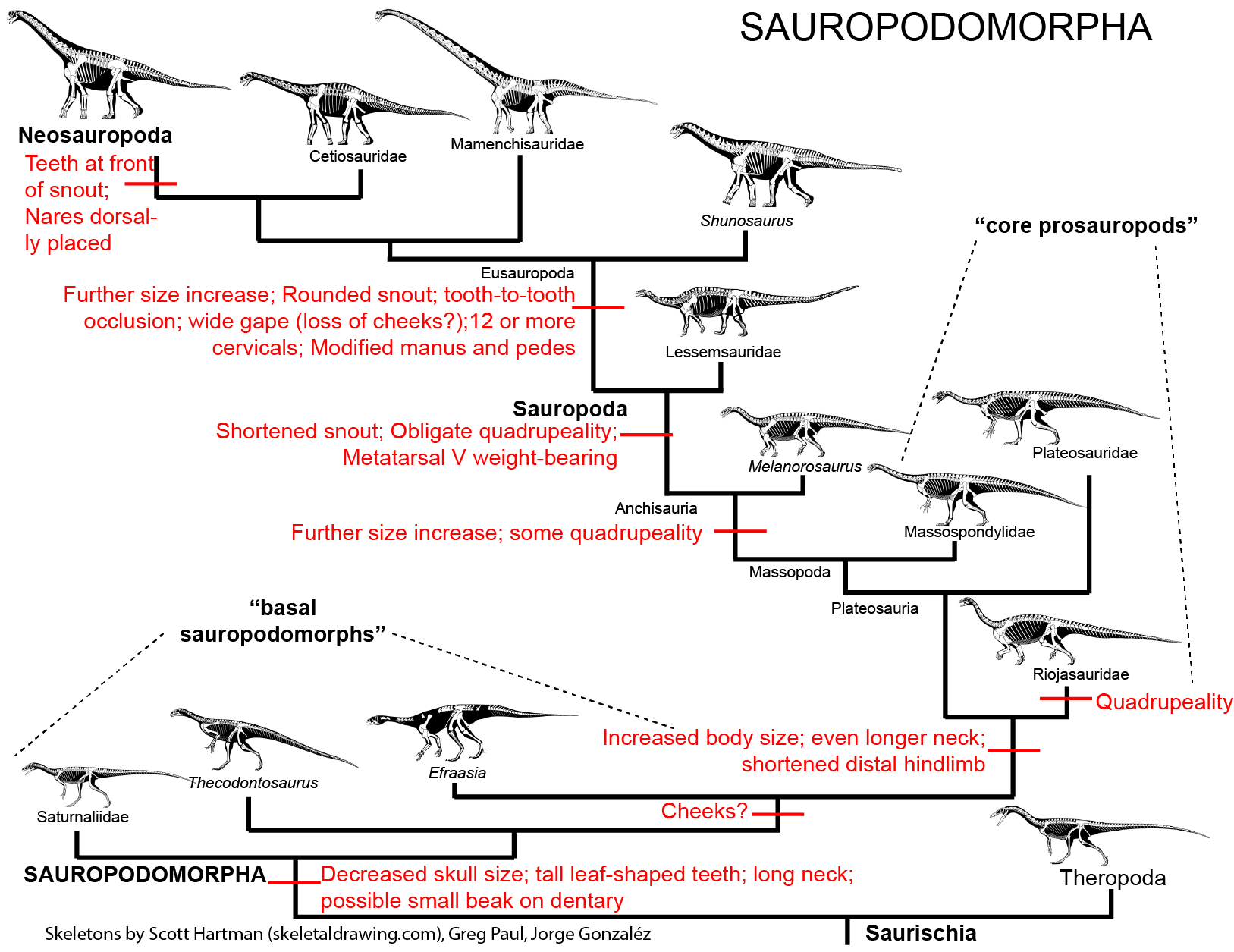
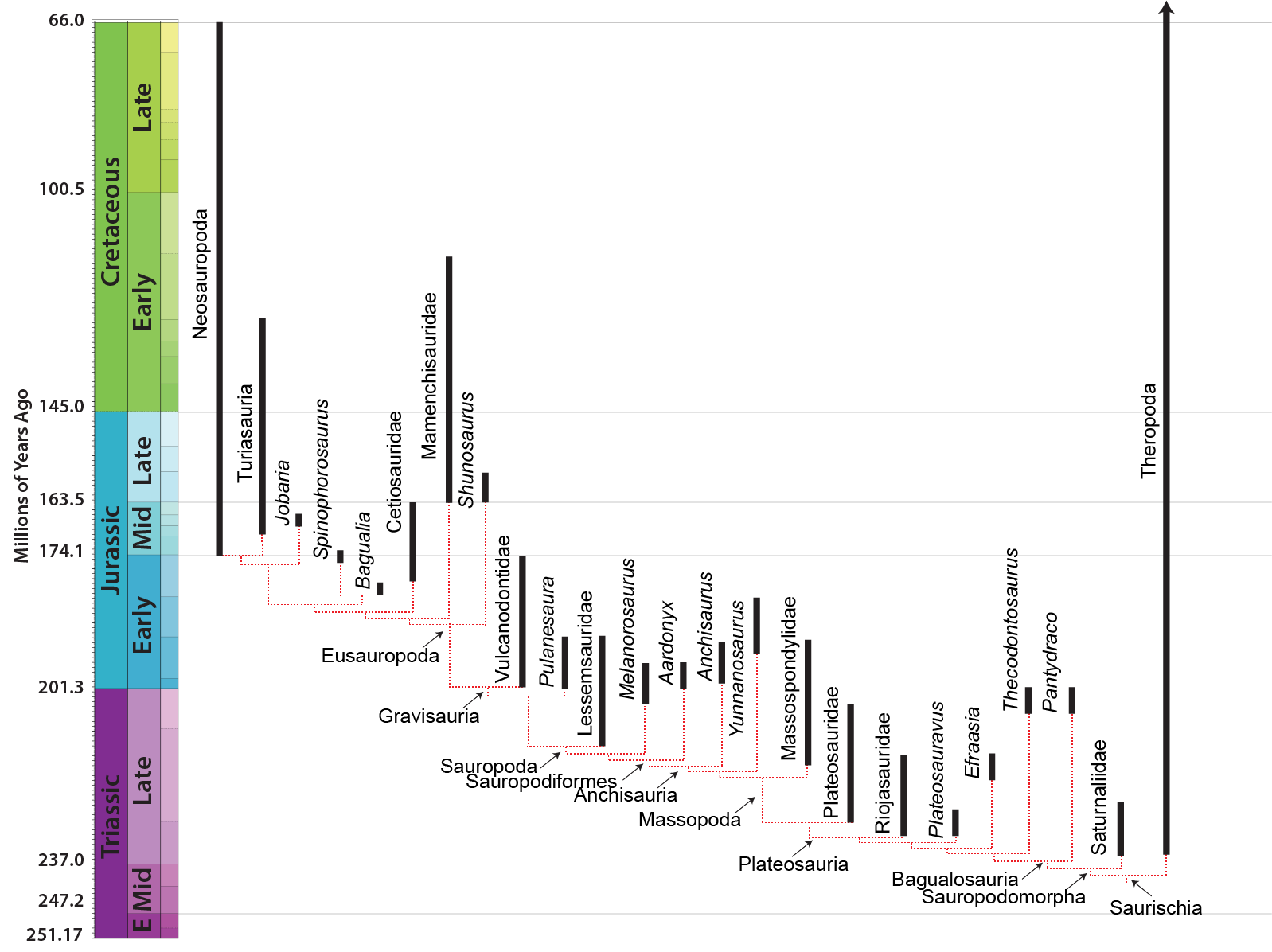
MAJOR GROUPS OF SAUROPODOMORPHS
The derived sauropodomorphs are the Sauropoda (lizard feet), giant obligate quadrupeds that include the largest land animals of all time. Primitive sauropodomorphs (i.e., sauropodomorphs other than sauropods) were traditionally grouped together as "Prosauropoda". Modern studies differ as to whether all prosauropods form a paraphyletic grade (with no major prosauropod groups more closely related to each other than to sauropods), while others recognize a monophyletic "core Prosauropoda" composed of plateosaurids, riojasaurids, massospondylids, etc. This course follows the first (paraphyletic grade) hypothesis (see cladogram and phylogeny above).
BASAL SAUROPODOMORPHS
Specializations of the most primitive sauropodomorphs relative to other saurischians include:
These forms were small (~1-2.5 m long), bipedal plant eaters. Their long necks would allow them to feed on higher leaves than most of their contemporaries. Recent work suggest a paraphyletic series of basalmost sauropodomorphs. The most completely known of these are Panphagia and Eoraptor of the Late Triassic of Argentina. (Eoraptor was long thought to be a basal theropod dinosaur, hence its name.) Newly discovered Chromogisaurus lived alongside them. Additional basal sauropodomorphs include Saturnalia and possibly Guaibasaurus of the Late Triassic of Brazil (Guaibasaurus is considered to be a basal theropod in some analyses), as well as other Late Triassic taxa around the world. The teeth of these basalmost forms are less phyllodont than the more derived taxa, and some teeth approach the ziphodont (blade-like) teeth of theropods in shapes: consequently, the earliest sauropodomorphs were almost certainly omniovres rather than strict herbivores. (In fact, that is the etymology for the name of Panphagia, "all eater" or "omnivore".)
Slightly more derived basal sauropodomorphs Thecodontosaurus, Asylosaurus and Pantydraco of Late Triassic England; Efraasia of Late Triassic Germany; and Plateosauravus of Late Triassic South Africa. Mussaurus of the Late Triassic of Argentina may be one of these basal forms, or may be more advanced (a "core prosauropod" or even a near-sauropod).
Like most saurischians, basal sauropodomorphs and core prosauropods had a "wrap around overbite": the toothrow of the mandible fit entirely within the tooth row of the upper jaw. Efraasia and later prosauropods (and basal sauropods) seem to have had a small fleshy cheek similar to (but smaller than) those interpreted for ornithischians. Less convincingly, some evidence suggests a small keratinous beak in front of the dentary (and possibly premaxilla) of basal sauropodomorphs and/or core prosauropods. The diets of basal sauropodomorphs would have included at least some plants, but these may have been more omnivorous than later, more derived sauropodomorphs.
CORE PROSAUROPODS
Plateosauridae (including Late Triassic European Plateosaurus),
Riojasauridae (including Late Triassic Argentine Riojasaurus), and Massospondylidae (including Early Jurassic South African
Massospondylus,
Argentine Adeopapposaurus,
Antarctic Glacialisaurus, North American Seitaad,
and Chinese Lufengosaurus): considered by some to form a
monophyletic "Prosauropoda" and by others a paraphyletic series relative to melanorosaurids and sauropods. Core prosauropods were the dominant
group of herbivores worldwide in the later Late Triassic and Early Jurassic: in some communities they were up to 95% of the known biomass. They represent
the first time dinosaurs dominated a major way of life. (They are the only common dinosaurs in Late Triassic Europe, for example.) Traditionally
interpreted as facultative bipeds, new studies of their forelimbs (especially forearms and wrists) show that they were not used for locomotion in most
of these and so these dinosaurs were obligate bipeds, although slowly built ones. (In Riojasaurus, however, limb proportions show that they were
almost certainly primarily quadrupedal.) Like basal sauropodomorphs, the core prosauropods retained the ancestral dinosaurian grasping hand. Their
distal hindlimb (tibia, fibula, and metatarsus) was shorter than in other saurischians (a trait they passed on to Sauropoda).
Larger than the basalmost sauropodomorphs, most core prosauropods range from 3 to 8 m longer (although some grew larger, up to or more than 10 m). Their necks were proportionately even longer than in their ancestors. They have been found in many environments: from swamps to deserts.
Although very common in the Late Triassic and Early Jurassic, no core prosauropods are known after the end of the Early Jurassic. They may have "evolved themselves into extinction", by being outcompeted by their specialized descendants: the sauropods.
"NEAR-SAUROPODS" AND THE PROSAUROPOD-SAUROPOD TRANSITION
The most advanced prosauropods (Anchisaurus, Yunnanosaurus, Lamplughsaura
of India, Leonerasaurus of Argentina, and Melanorosauridae (such as Late Triassic South African
Melanorosaurus) are considered basal sauropods by some workers. For the purpose of this course, we'll restrict "Sauropoda" to Saltasaurus
and everything sharing a more recent common ancestor with it than with Melanorosaurus, and use the unofficial name "near-sauropods" for those
sauropodomorphs phylogenetically intermediate between Massospondylus and Sauropoda.
Like sauropods, the near-sauropods were typically larger than the more primitive sauropodomorphs: in this case, some 10-13 m long or more as adults and perhaps 5 tons. (Anchisaurus [sometimes called Ammosaurus] is a tiny excpetion: only as big as the basal sauropodomorphs.) Recently discovered Aardonyx of the Early Jurassic of South Africa is the last branch of Sauropodomorpha which was an clearly an obligate biped. Most near-sauropods were at best facultative bipeds, and some may have been obligate quadrupeds. Most add another sacral vertebrae (total of four).
The evolution of sauropodomorphs shows a correlated progression towards larger body size, smaller heads, longer necks, and quadrupedality. This may have been related to two (not mutually exclusive) selective forces:
These transitional near-sauropod forms are Late Triassic-to-earliest Middle Jurassic in age.
SAUROPODA
Postcranially and in size (10-13 m length), basal sauropods (Late Triassic Antetonitrus,
Isanosaurus, Gongxianosaurus, and the Vulcanodontidae) do not differ much from
melanorosaurids and other near-sauropods. But early sauropod skulls have been transformed:
In vulcanodonts and the more derived eusauropods (together the Gravisauria) there are four or more sacrals, and there is a transformation of the jaws and teeth:
Early sauropods are known from the southern continents (Gondwana) in the Late Triassic, and only show up in northern continents in the Early Jurassic. They seem to have been only rare components in dinosaurian communities until the Middle Jurassic.
EUSAUROPODA
The sauropods of the Middle Jurassic onward comprise the Eusauropoda (true sauropod). Eusauropods differ from their ancestors in a number of ways:
There has been an ongoing debate over the position of the necks of eusauropods (well, long-necked sauropodomorphs in general, but especially for these guys). Computer studies of possible ranges of motion in sauropod necks suggested that they were basically horizontal. However, recent work on a diversity of living animals shows an upright curved neck in nearly all amniotes, so it is quite likely that a raised neck was the natural position for sauropods.
Recently discovered Spinophorosaurus of the Middle Jurassic of Niger is the most completely known primitive sauropod. It might be a eusauropod, or it might be the sister taxon to Eusauropoda. Its tail (like that of Middle Jurassic Chinese Shunosaurus) ends in a tail club: in fact, that of Spinophorosaurus is convergent in form on stegosaurian thagomizers.
Eusauropods are the dominant herbivores worldwide in the Middle and Late Jurassic, and in at least some parts of the world all the way until the end of the Cretaceous. The best and most completely known early eusauropod is Middle Jurassic Shunosaurus of China. Jobaria of the Middle Jurassic (once thought to be Early Cretaceous) of Africa has sometimes been considered a primitive member of Macronaria, but other analyses suggest it was a typical member of this early eusauropod radiation.
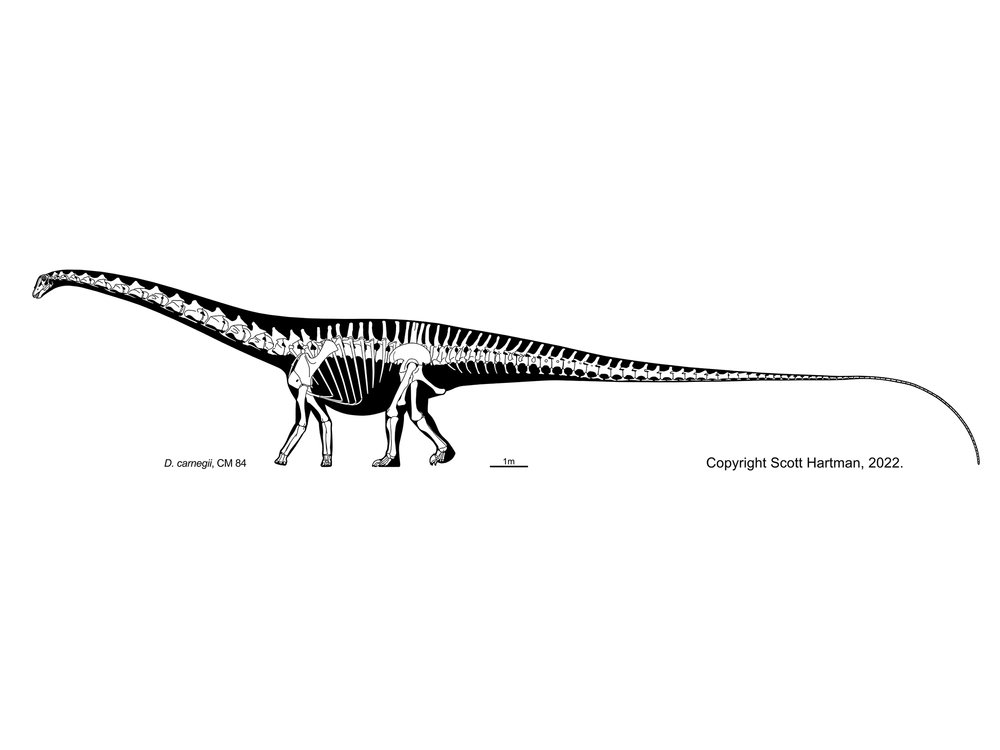
Several groups of primitive eusauropods are known. Some recent work suggests that many form a clade Cetiosauridae, named after Cetiosaurus of the Middle Jurassic of England. Other cetiosaurids include Patagosaurus of the Middle Jurassic of South America and Barapasaurus of the Early Jurassic of India. However, other cladistic studies suggest that "cetiosaurids" are a paraphyletic series of primitive eusauropods (and that even "Cetiosaurus" is not monophyletic.)
Different clades of eusauropods show different specializations towards reaching higher in trees. Several Middle-Late Jurassic Asian sauropods evolved extra cervical vertebrae which were proportionately very long. Some of these sauropods (including Middle Jurassic Omeisaurus and Late Jurassic Mamenchisaurus) seem to form a clade, Mamenchisauridae. Some mamenchisaurids seemed to have reached truly tremendous size.
A recenly discovered but still poorly known group of eusauropods of the Late Jurassic of Europe may form a monophyletic group Turiasauria. Some of these "turiasaurs" evolved into tremendously large forms (rivalling various giant neosauropods).
NEOSAUROPODA
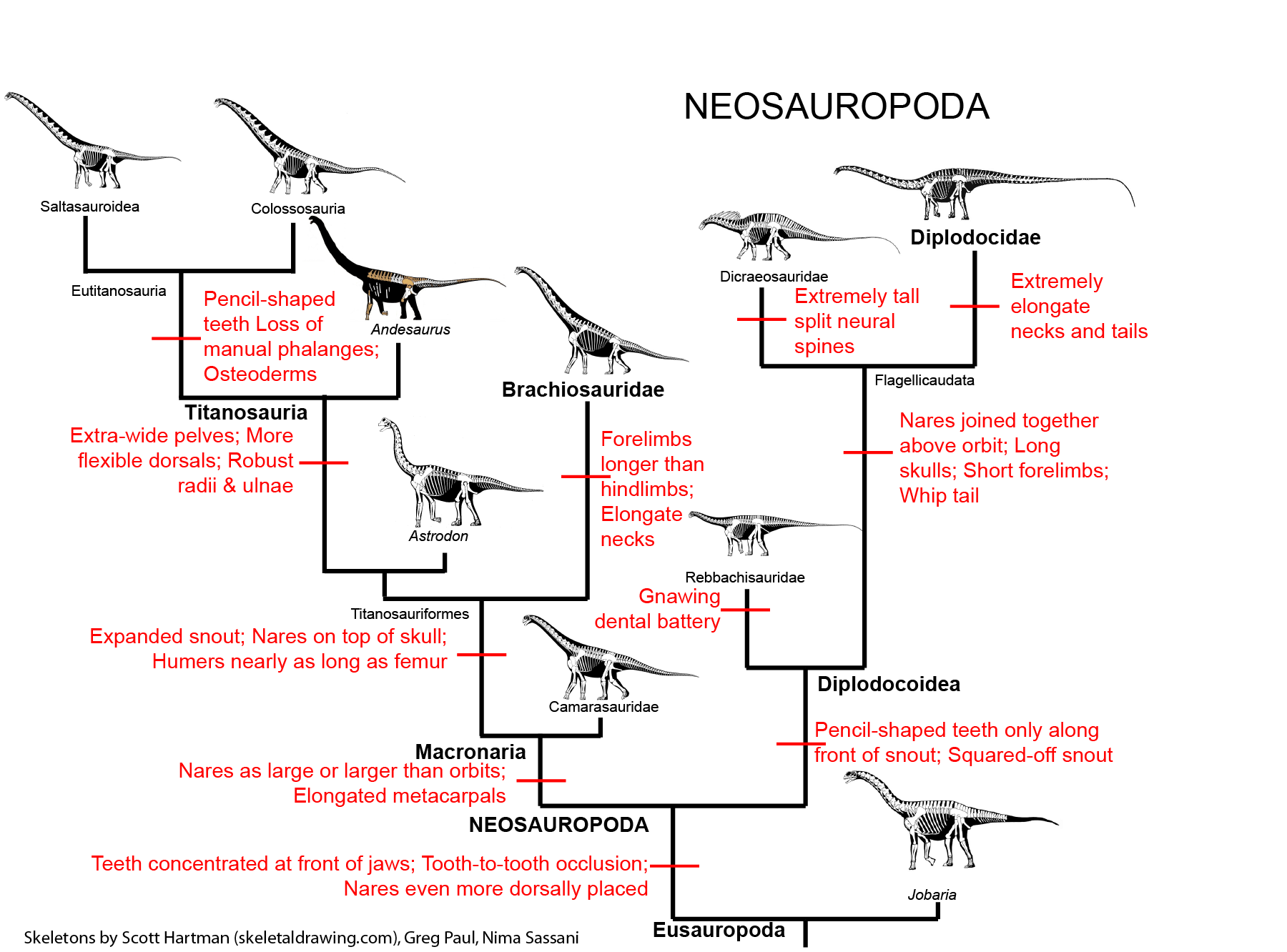
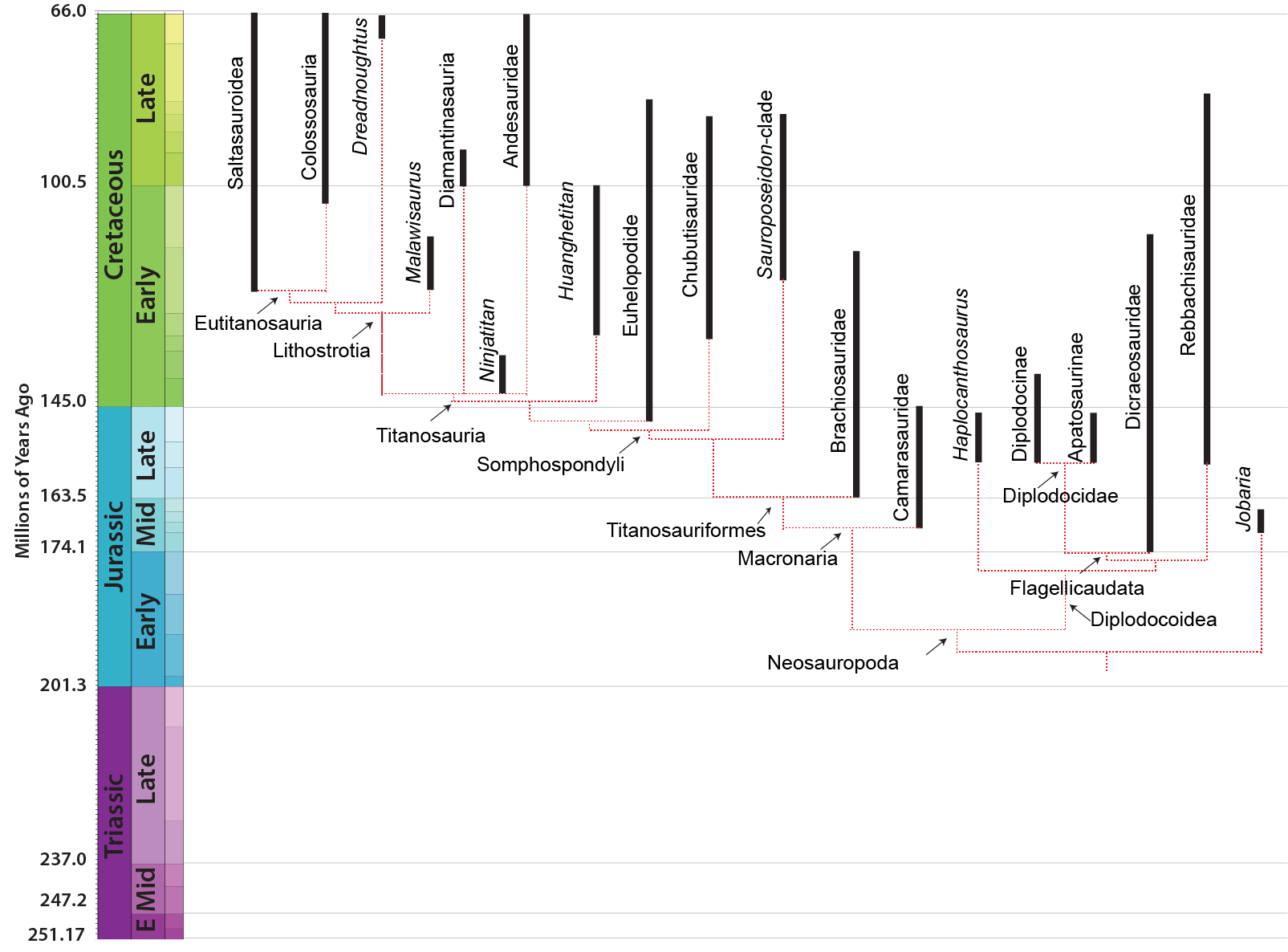
The dominant group of post-Middle Jurassic sauropods is the Neosauropoda (new sauropods). Their transformations from primitive eusauropods are:
Neosauropods have two major subdivisions: Diplodocoidea and Macronaria.
DIPLODOCOIDEA
This group differs from their ancestors by a number of traits:
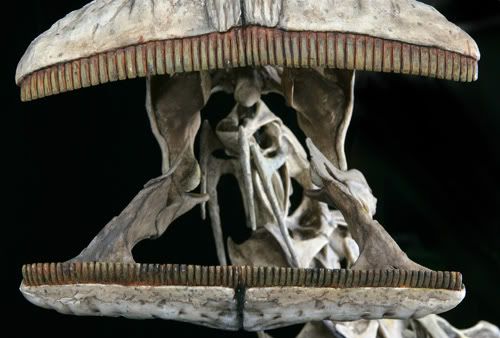
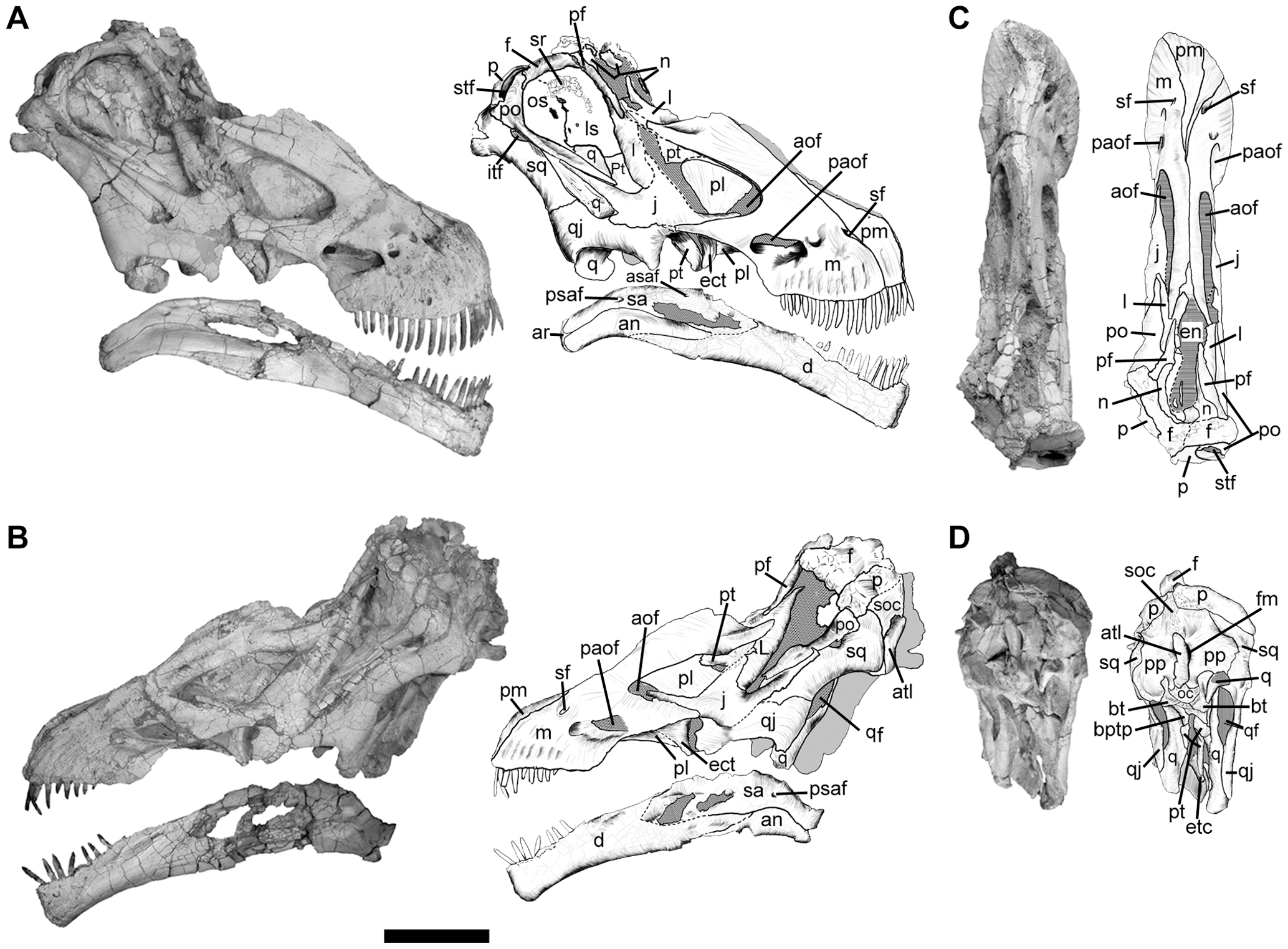
The first major branch of diplodocoids is the truly bizarre Rebbachisauridae, currently known from the Cretaceous, mostly from Gondwana. They are best known from Nigersaurus from the late Early Cretaceous of northern Africa. At least Nigersaurus was "little" (as sauropods go, that is: only the size of an Indian elephant!). Their skulls are highly transformed, most importantly with:
Unlike the slicing dental battery of ceratopsids or the grinding dental battery of hadrosaurids, the rebbachisaurid dental battery is at the front end of the jaw. The rate of replacement of the teeth was very high (one tooth/month; about twice as fast as hadrosaurids).
Other rebbachisaurids include Rebbachisaurus of northern Africa and Limaysaurus, Rayososaurus, Cathartesaura, and Nopscaspondylus of South America, and Histriasaurus and Demandasaurus of Europe.
The remaining diplodocoids are the Flagellicaudata (whip tails). These form a major part of the sauropod communities of the Late Jurassic and early Early Cretaceous. Their specializations include:
There are two major divisions of the flagellicaudates. The Dicraeosauridae were smaller (only 10 m or less), and had:
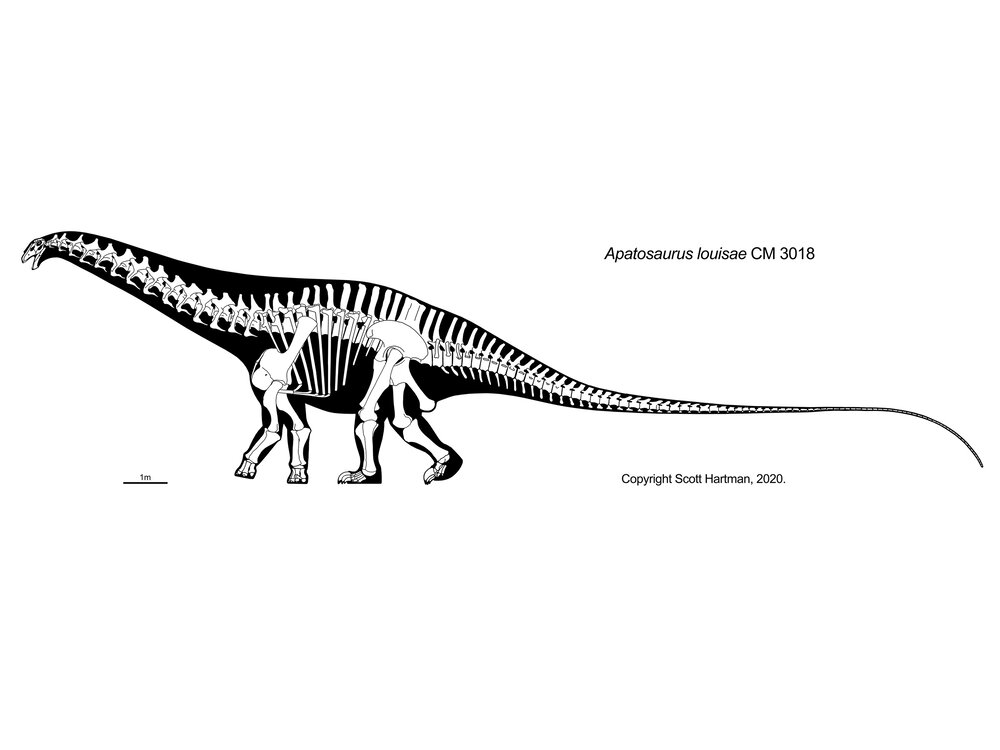
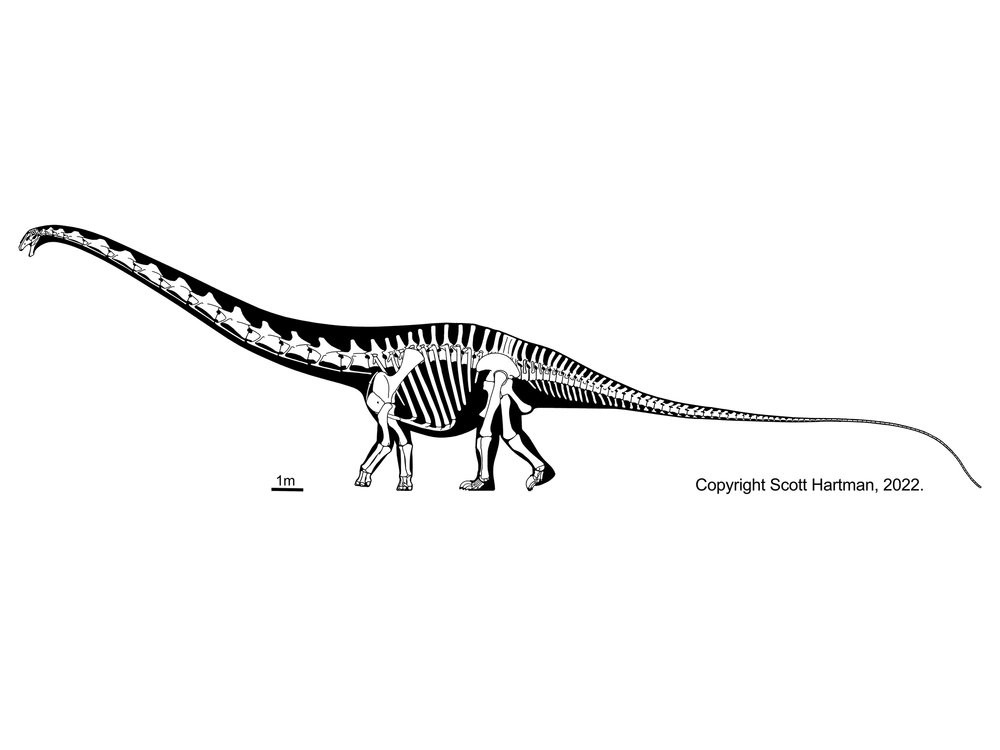
More famous are the Diplodocidae. These are very large sauropods (over 30 m in massively-built Apatosaurus, and more slender Diplodocus [the individual formerly considered Seismosaurus], Supersaurus, and Amphicoelias). They are characterized by the following specializations:
Diplodocids (and other diplodocoids?) may have been able to rear up on their hindlimbs to feed even higher in trees than they could using their long necks. Alternatively, they may have been low-browsing/grazing specialists: we'll come back to this question in a later lecture. At present, true diplodocids are known only from the Late Jurassic. In western North America they were tremendously diverse: at least six genera were present at one time.
MACRONARIA
The other major branch of neosauropods is the Macronaria (big noses). They range from the Late Jurassic to the very end of
the Cretaceous. Their changes from ancestral sauropods include:
Primitive macronarians include Camarasaurus of the Late Jurassic of western North America (and possibly elsewhere); Tehuelchesaurus of Late Jurassic Argentina; and others.
(However, a note for the future: some big on-going phylogenetic analyses threaten to shake up the family tree. It may be that Camarasaurus will fall outside of a clade comprised of diplodocoids, brachiosaurids and their kin, and titanosaurs. Stay tuned.)
The more advanced macronarians (the Titanosauriformes) are specialized by:
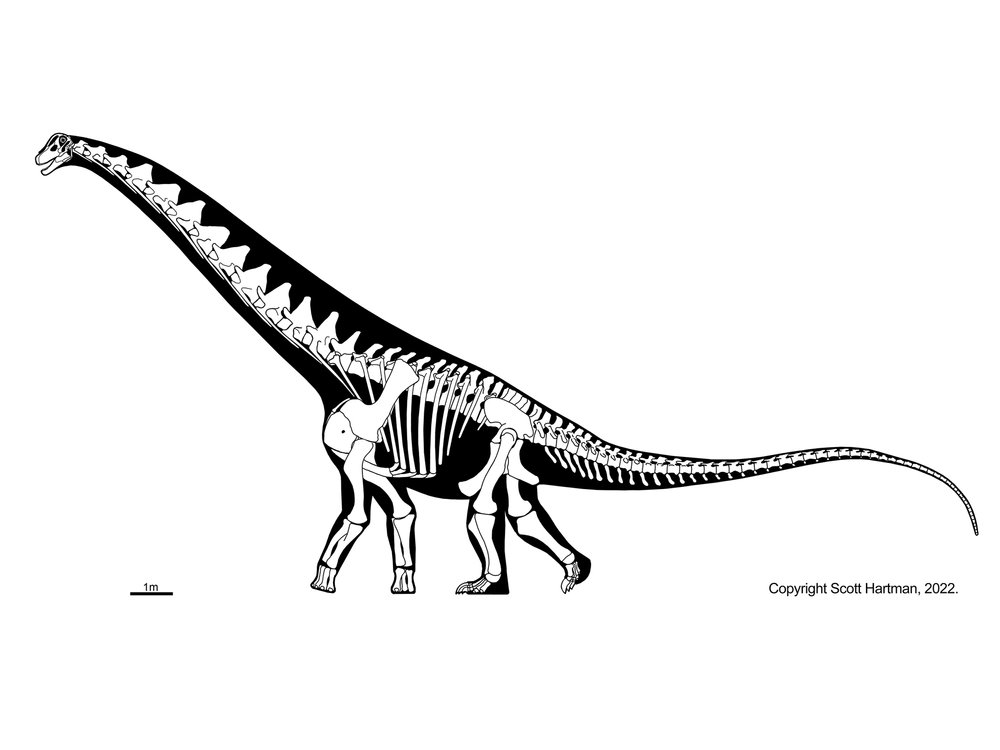
Astrodon (State Dinosaur of Maryland; Early Cretaceous of Maryland, and possibly Texas) is a titanosauriform, as is as is gigantic Sauroposeidon of Early Cretaceous western North America. Three major clades of titanosauriform include the Brachiosauridae, the Euhelopodidae, and Titanosauria.
Brachiosauridae (Middle Jurassic to Early Cretaceous) are very distinctive. They are characterized by:
The result is that brachiosaurids were built "up hill", making them some of the tallest animals that ever lived. Best known are Late Jurassic Brachiosaurus of North America, Giraffatitan of Africa, Lusotitan and Europasaurus of the Late Jurassic of Europe, and Early Cretaceous Abydosaurus, Venenosaaurus, and Cedarosaurus of western North America. Astrodon and Sauroposeidon are sometimes considered to be brachiosaurids. Alternatively, some studies find the "brachiosaurids" as a paraphyletic series with regards to euhelopodids + titanosaurs.
The Euhelopodidae is a clade of ultra-long-necked Cretaceous titanosauriforms. At present all known members are from Asia. They include (among others) Early Cretaceous Euhelopus, Qiaowanlong, Phuwiangosaurus, and early Late Cretaceous Erketu. Daxiatitan and Ruyangosaurus of the early Late Cretaceous of China may both be euhelopodids: the latter is one of the contenders of "largest dinosaurs of all time", with a 2.35 m (7.7 foot) tall femur. (However, some traits of the vertebrae of Ruyangosaurus suggest it is a titanosaur, close to comparably-gigantic Puertasaurus.
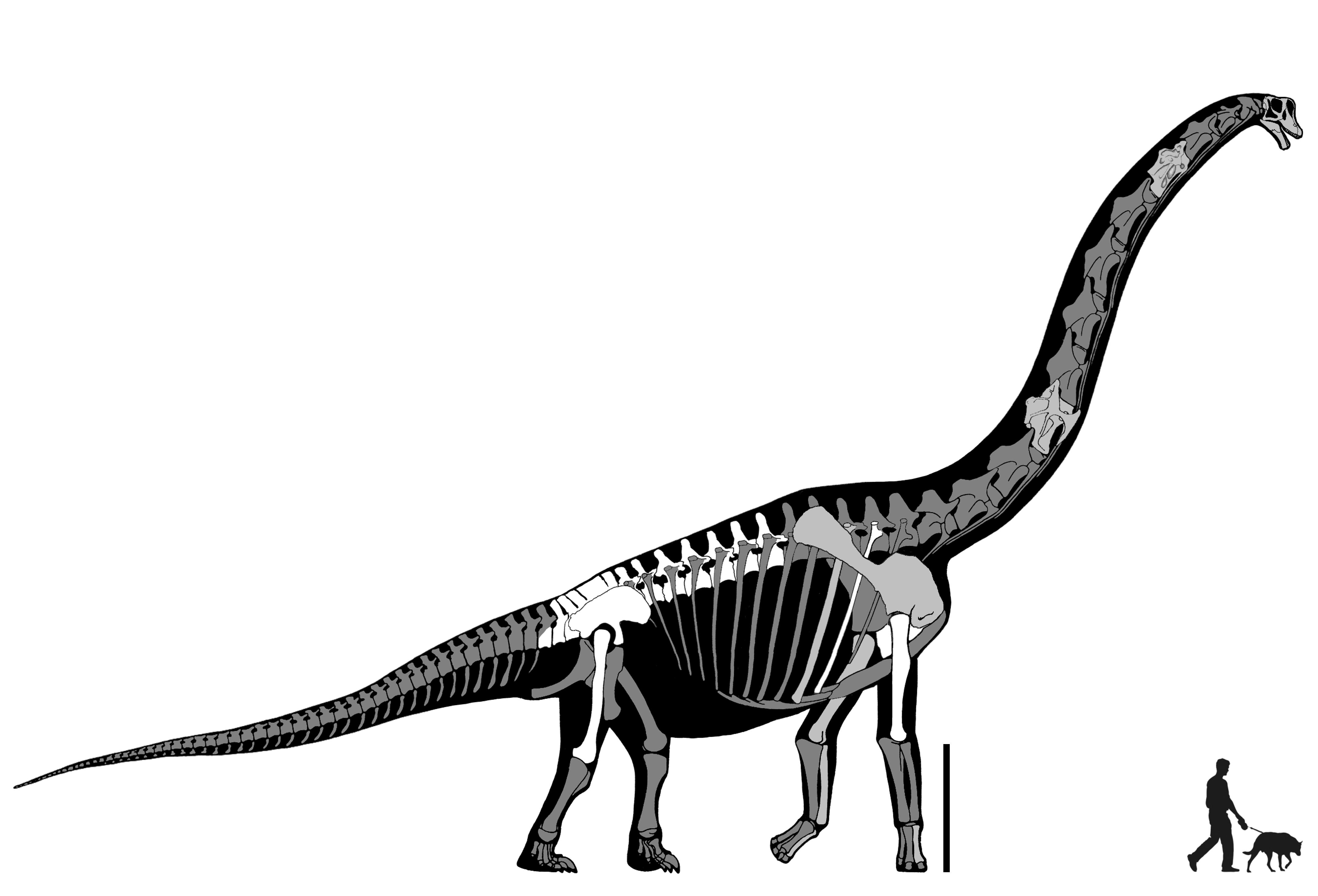
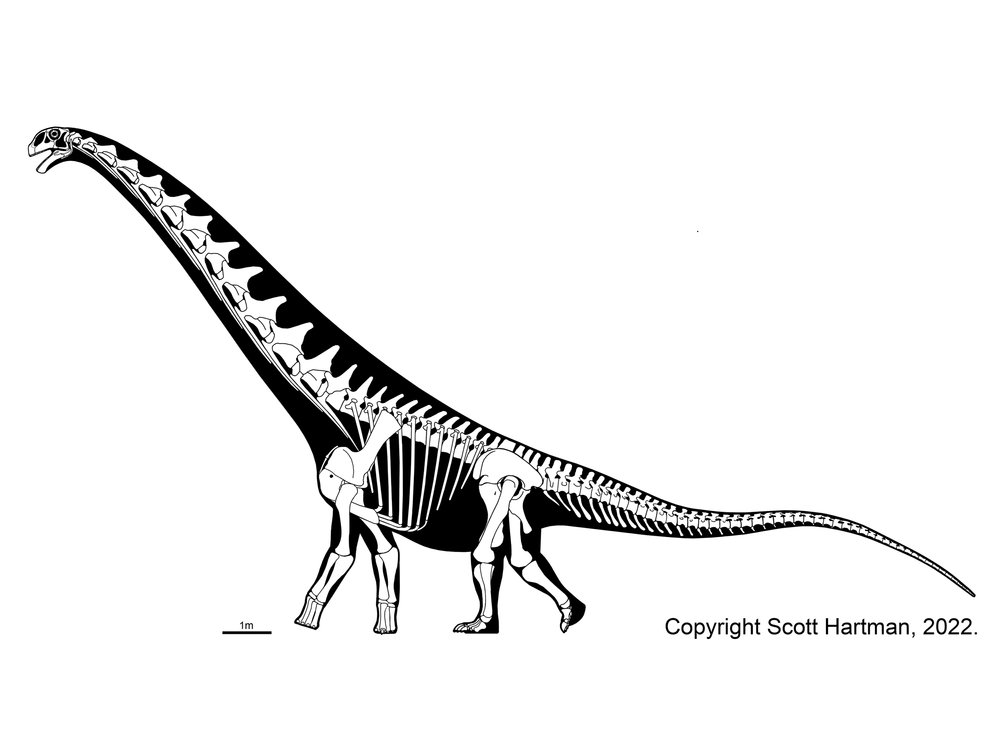
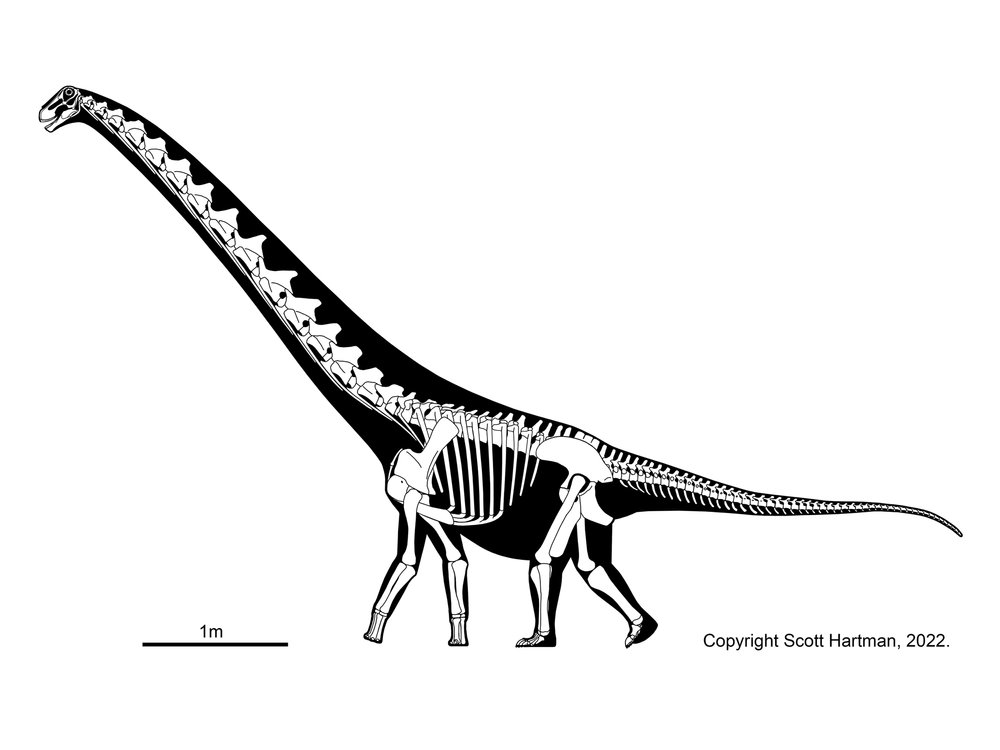
Titanosauria (Cretaceous) were the dominant group of Cretaceous sauropods in most parts of the world. They were very diverse. The different groups share:
In addition to some primitive taxa, the titanosaurs include a more derived clade Eutitanosauria, which is further modified by the evolution of:
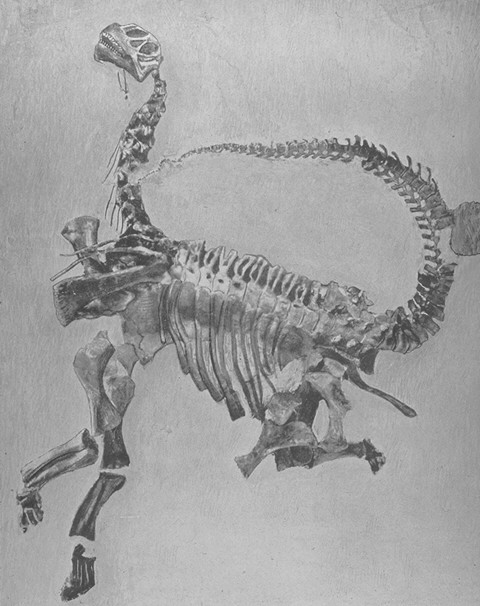
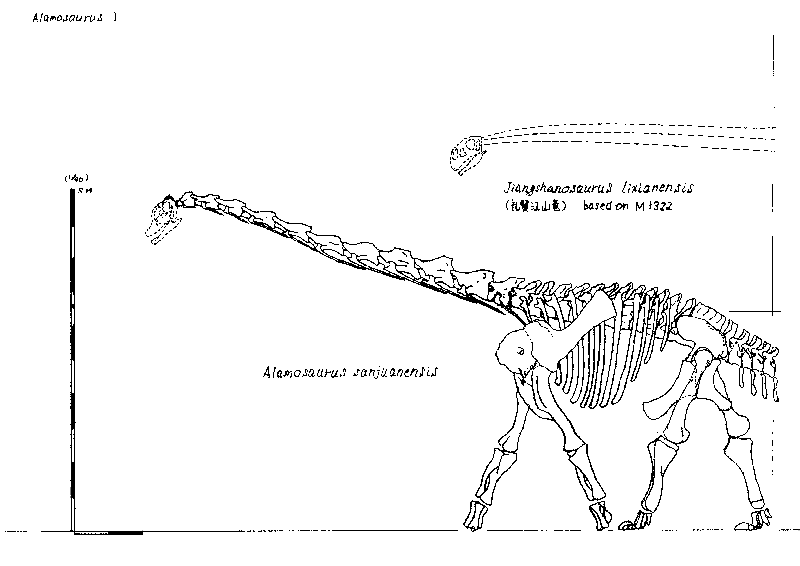
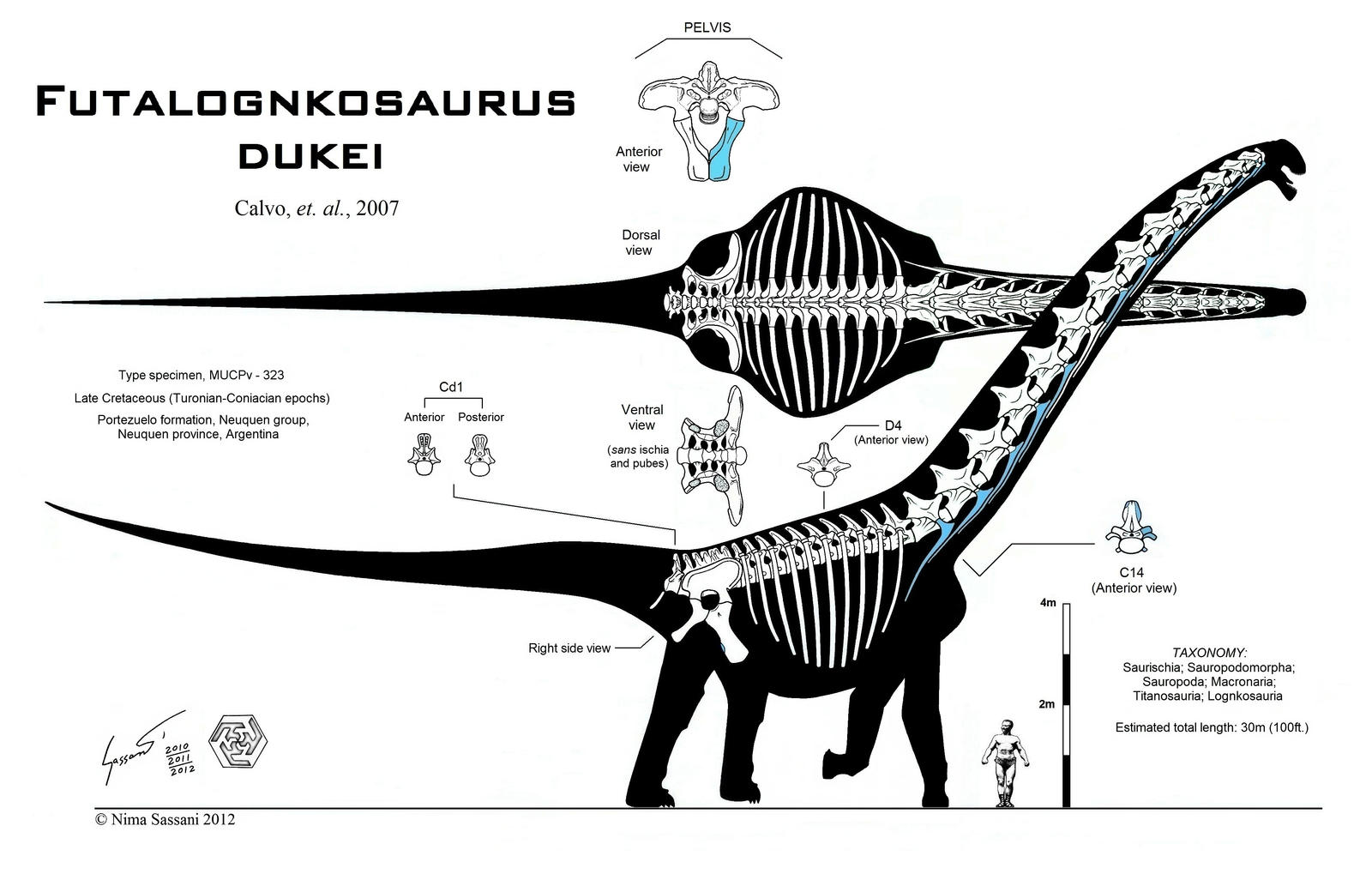
Relationships among the eutitanosaurs are still being worked out. What is presented here is likely NOT going to be strongly supported in more comprehensive studies, but works as a provisional scheme. Among the eutitanosaur groups are the enormous heavily built Argyrosauridae, the long-necked the long-necked Lognkosauria (including Futalognkosaurus, one of the largest dinosaur known from relatively complete material), the somewhat brachiosaurid-like Antarctosauridae (including 90-100 ton Argentinosaurus, perhaps the largest dinosaur of all, and giant Alamosaurus (the last North American sauropod)), and the armored Lithostrotia. The latter were relatively smaller than other titanosaurs, and include the Saltasauridae and the broad-snouted Nemegtosauridae (including Early Cretaceous Brazilian Tapuiasaurus, known from a very well-preserved skull.)
Titanosaurs are the main group of herbivorous dinosaur in the southern continents (and parts of Europe) until the end of the Cretaceous, and were found on all continents. Titanosaurs include some truly tremendous forms: Futalognkosaurus, Argentinosaurus, Puertasaurus, and Alamosaurus all seem to reach just about the same size: masses of perhaps 100 tons or so, and lengths much more than 30 m. These dinosaurs are still smaller (less massive) than the modern blue whale, but otherwise exceed any living species of animal in mass. We will explore later on HOW it was that dinosaurs could be able to reach this supremely gigantic sizes.
EVOLUTIONARY PATTERNS IN SAUROPODOMORPHA
Feeding adaptation transformations:
Gigantism:
Locomotory changes:
Heterochrony and the origin of Sauropoda:
Niche partitioning:
Many dinosaur communities had multiple sauropod species at the same time. How could they support this diversity? Niche partitioning: each clade
(and even each species) specialized in feeding in a slightly different manner. For example, in Late Jurassic communities there were ground-feeding
dicraeosaurids, mid-level feeding primitive eusauropods and primitive macronarians, high feeding brachiosaurids, mamenchisaurids, and diplodocoids
(which could feed exceptionally high if they could actually rear on their hindlegs)
To Next Lecture.
To Previous Lecture.
To Syllabus.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}