
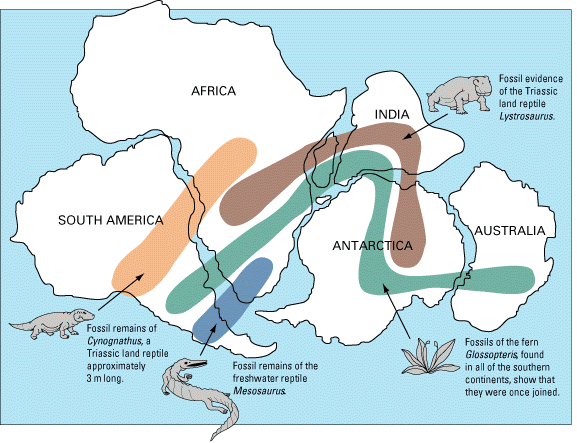
Some early indication that continents may have moved in past:
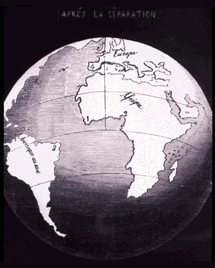
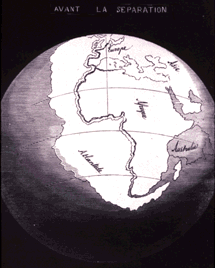
Alfred Wegener proposed model of continental drift (in 1915):
However, big flaws in continental drift:
Idea was considered unlikely by many geologists.
In 1940s through 1960s, new information. Harry Hess and colleagues discovered that fossil, radiometric, and magnetic dates showed that ocean basins were not ancient, but that they got younger closer to mid-ocean ridges, where new rock was forming. Hess called this "sea-floor spreading".
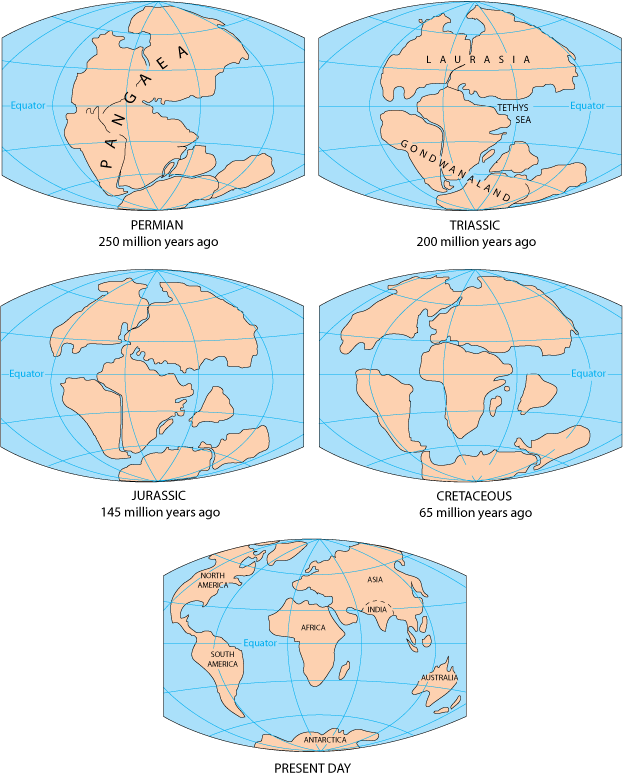
In 1960s, the models of continental drift and sea-floor spreading were combined by John Tuzo Wilson and colleagues to form plate tectonics.
Heat from Earth's core moves plates, forming mountain ranges at subduction and collision boundaries. Weather erodes uplifting mountains, wind and water and ice transports sediment to depositional environments. Over time, material becomes buried.
Plates wander over Earth's surface, so continents move from tropics to poles or back. Also, action of mid-ocean ridges causes sea levels to rise up (flooding continents) or lower (draining continents). (Current situation is very low sea level).
Big change from the 1960s-1970s model: now recognize there are LOTS of little plates (terranes) rather than just a few big plates. See here for a detailed look at the changing shape of Earth's surface for the last 600 million years; and here for a close-up on North American paleogeography.
Plate tectonics ultimately drives geology:
First up, some definitions:
How does energy go from the non-living environment into the living world? One major way is photosynthesis. Here is the process greatly simplified:
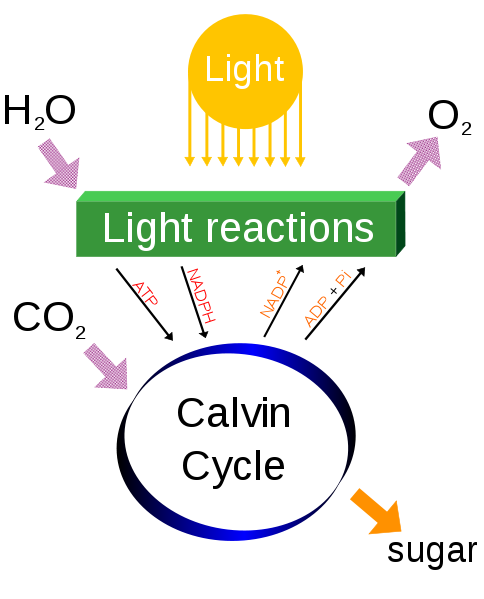
And here is the photosynthesis equation, simplified:
6CO2 + 6H2O + Sunlight yields C6H12O6 + 6O2
Or, in other words, photosynthesizers (plants and algae) take in water and carbon dioxide and using sunlight convert these into sugars and waste oxygen gas. Photosynthesizers are the major form of autotroph: organisms that don't eat other organisms.
In contrast, most other life forms (animals, fungi, most "protists", etc.) are heterotrophs: we get nutrients from digesting other life forms, primarily via the aerobic respiration equation:
C6H12O6 + 6O2 yields 6CO2 + 6H2O + Energy
(That is, food (glucose) plus oxygen yields waste carbon dioxide and waste water, plus energy).
In this reaction, we take in sugars and oxygen and release waste water and carbon dioxide.
So plants, algae, and other photosynthesizes are basically made up of air (and a little bit of other matter drawn from soil and/or water), while animals and other heterotrophs are made of food (ultimately plants) and air. This most important aspect of ecology is thus trophic relationships: who eats whom. These mark out relationships in a food chain of organisms that eat the organisms below them. However, since in nearly every case any species eats more than one type of food, trophic relationships are better displayed as a food web, comprised of many interlocking food chains.
The trophic level is the position an organism occupies of a food chain. Producers do not consume other animals: in terrestrial ecosystems, these are plants (which "eat" sunlight). Decomposers break down the bodies of any kind of organisms. Consumers are those that eat other organisms: 1st order consumers are herbivores (plant-eaters); 2nd order consumers eat 1st order consumers; 3rd order consumers eat 2nd and 1st order consumers; and so on, up to the apex predator (consumers that eat lower order consumers but are not typically eaten by any species but themselves).
As you move up a food chain, there is less and less energy available. Plants receive much (but by no means all) of the energy of incoming sunlight; some of that energy becomes stored as their tissues, but the rest of it is used for them to run their bodies. Herbivores consume plants, and gain some (but not all) of that stored energy, some of which they used to build their own tissues but the majority being used for their own operation (running, breathing, pumping blood, etc., etc., etc.). Predators eat the herbivores, gaining some (but not all) the stored energy in the tissues of the herbivores, using some to build their own bodies, but the rest for their operations. Thus, the amount of energy available at each trophic level becomes less and less: this is described as the energy pyramid.
Assessing Trophic Relationships
One of the most important aspects of ecology (and paleoecology) is determining who ate whom: that is, reconstructing the food web. Sometimes
there is direct trace fossil evidence: for example, this humerus
of the Late Cretaceous Asian saurolophine Saurolophus: the arrows point to bite marks generated by a tyrannosaurid (almost certainly Tarbosaurus,
which is known from the same fauna).
However, sometimes we do not have such direct evidence. In these cases, we can make relatively reasonable assessements. For example, it is unlikely that small theropods habitually killed the largest herbivores in their environment (while the latter were adults, at least: babies would be fair game), and it is unlikely that there were species that were not eaten by SOMETHING.
In the case of the herbivores, however, it becomes somewhat more difficult. For one thing, it is often the case that the conditions that are best to preserve plant fossils (especially of leaves and fruits) are not good for preserving bone, and vice versa. So we often lack good knowledge of the flora that co-existed with any fauna. That said, we have the digested plant remains in corpolites as positive evidence that dinosaurs ate particular plant species (although referring a coprolite to a particular dinosaur species can be difficult.)
To Previous Lecture.
To Next Lecture.
To Syllabus.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}