
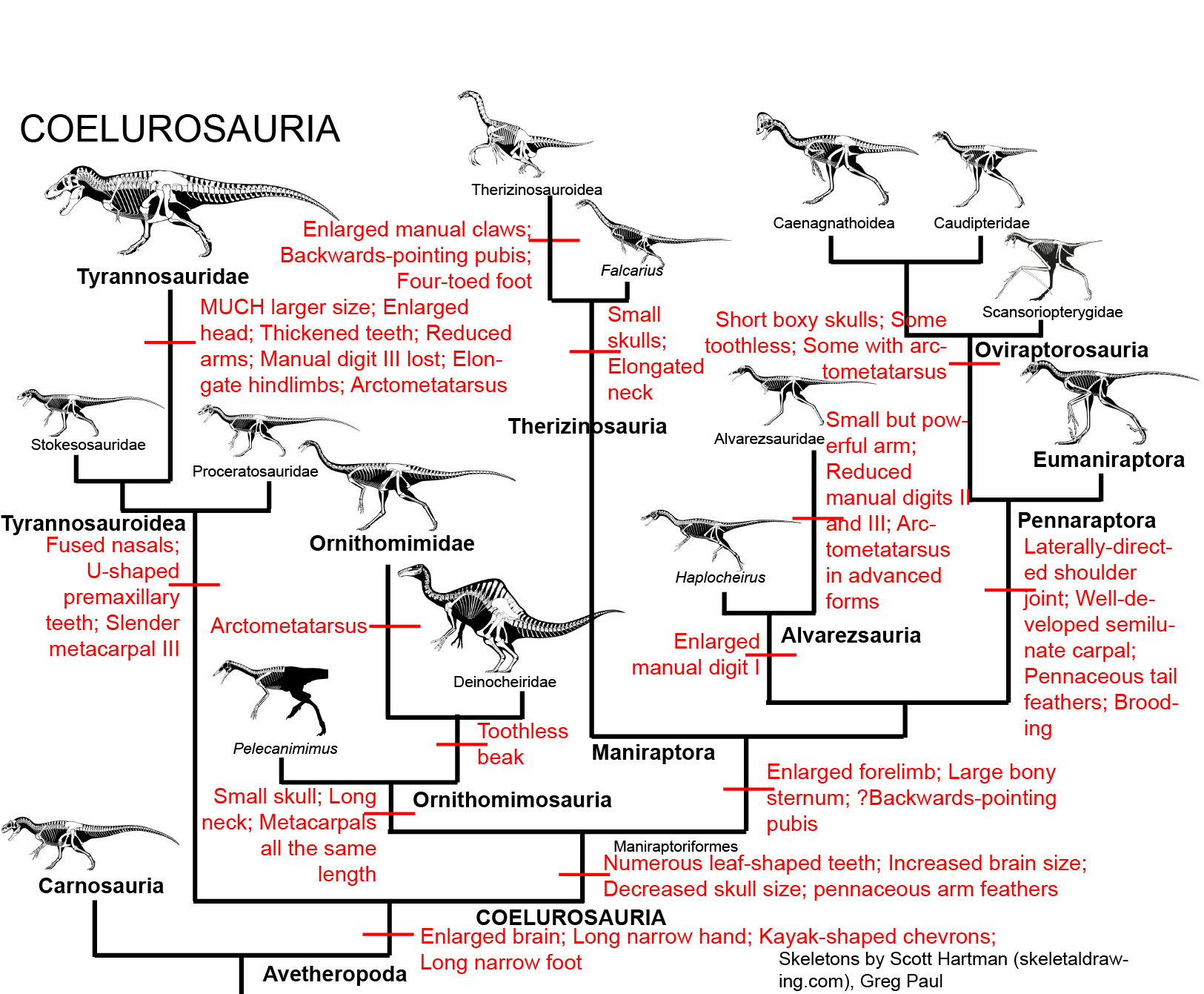
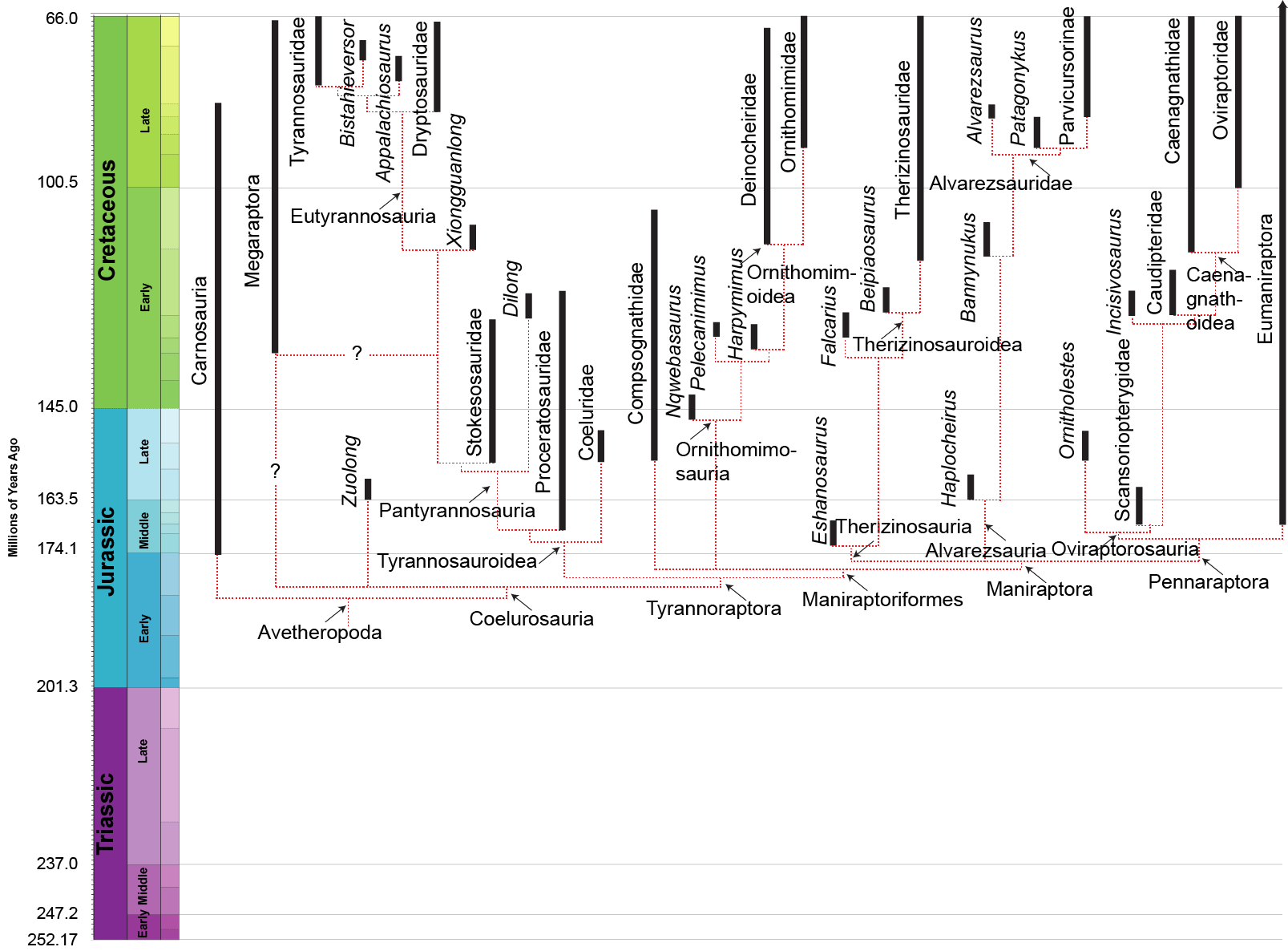
MAJOR GROUPS OF COELUROSAURS
Coelurosaurs are the sister group to Carnosauria within the avetheropods. The coelurosaurs differ from other theropods by possessing:
Additionally, where known, all coelurosaurs preserved in the appropriate-style sediment show at least some protofeathers or true feathers. However, as discussed previously, the discovery of the megalosauroid Sciurumimus pulls the origin of protofeathers much deeper into theropod (or dinosaur, or ornithodiran) history. At least some of the protofeathers of primitive coelurosaurs seem to be pennaceous (having a central shaft), rather than simply plumulose tufts.
Among the most primitive and oldest known coelurosaurs are the basal tyrannosauroids Proceratosaurus of the Middle Jurassic of England and Kileskus of Russia. Only the skull of the former, and skull, hand, and foot bones of the latter, are known at present. Other basal coelurosaurs include a trio of Late Jurassic western North American forms: Ornitholestes; Coelurus; and Tanycolagreus. It may be that one or more of these basal forms are actually basal tyrannosauroids. These and other early coelurosaurs were relatively small (2-4 m) slender animals with skulls full of small ziphodont teeth. Their narrow grasping hands suggest they adapted to catching small prey; their light build, slender limbs, and narrow dynamic stabilizing tail suggests relatively agile animals (useful both in chasing prey and in avoiding predators).
Newly described is a very late surviving basal coelurosaur, Bicentenaria of the mid-Cretaceous of Argentina.
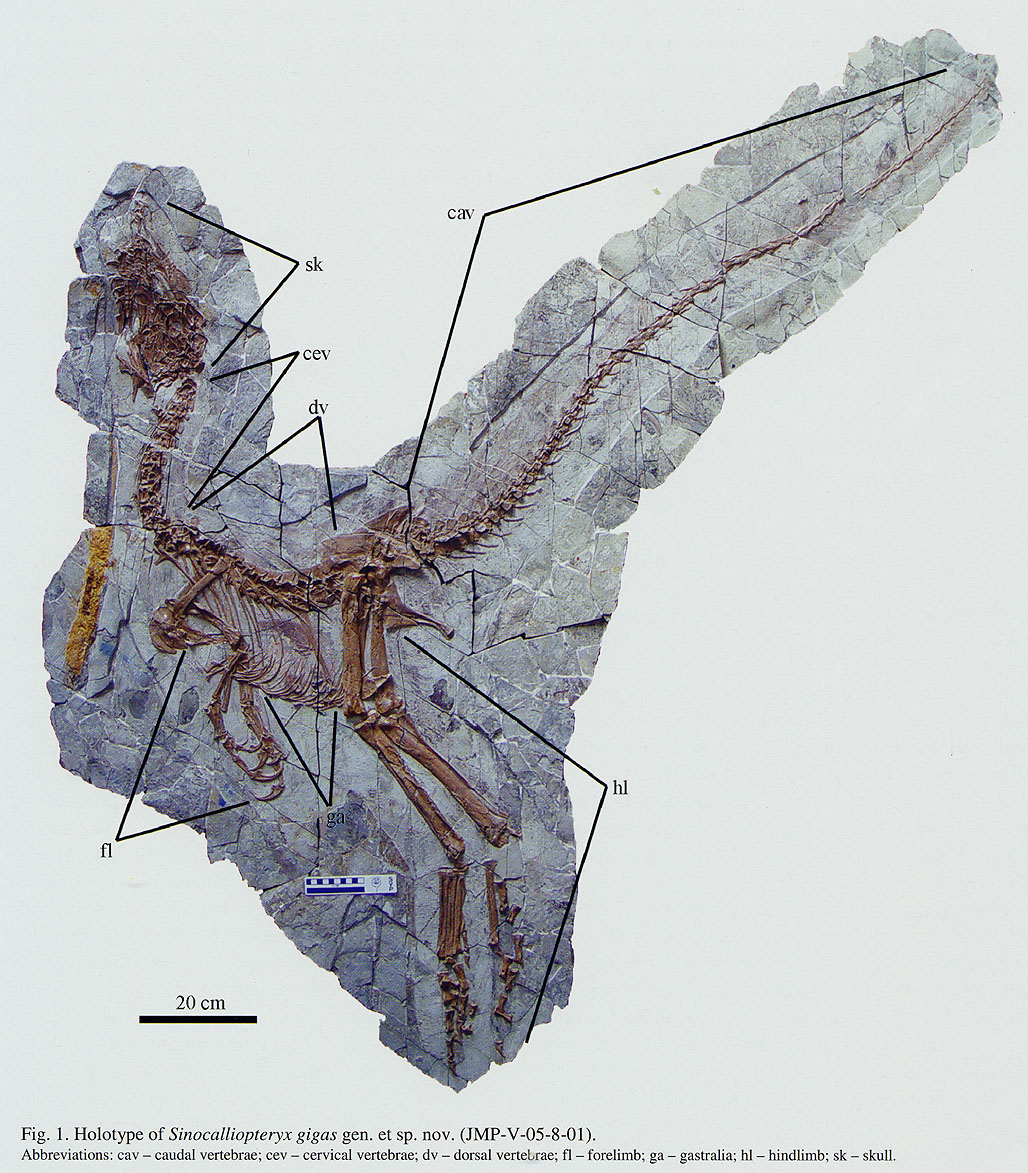
An important group of small primitive theropods is the Compsognathidae. This group ranges from the 1 m long Compsognathus of the Late Jurassic of Europe and Sinosauropteryx of the Early Cretaceous of China to 1.75 m long Huaxiagnathus of the Early Cretaceous of China to the "giant" Sinocalliopteryx of the Early Cretaceous of China at 2.5 m long. Compsognathids are also known from Early Cretaceous Europe and South America, and represented a minor radiation of small-bodied dinosaurs. Gut contents show that they ate lizards and small mammals.
TYRANNOSAUROIDEA
The most long-lived and ecologically significant group of primitive coelurosaurs was Tyrannosauroidea, the tyrant
dinosaurs. Best known from the later Late Cretaceous Asia and North American Tyrannosauridae, recent discoveries
reveal a long history of tyrant dinosaurs going back into the Middle Jurassic.
Basal tyrannosauroid specializations include:
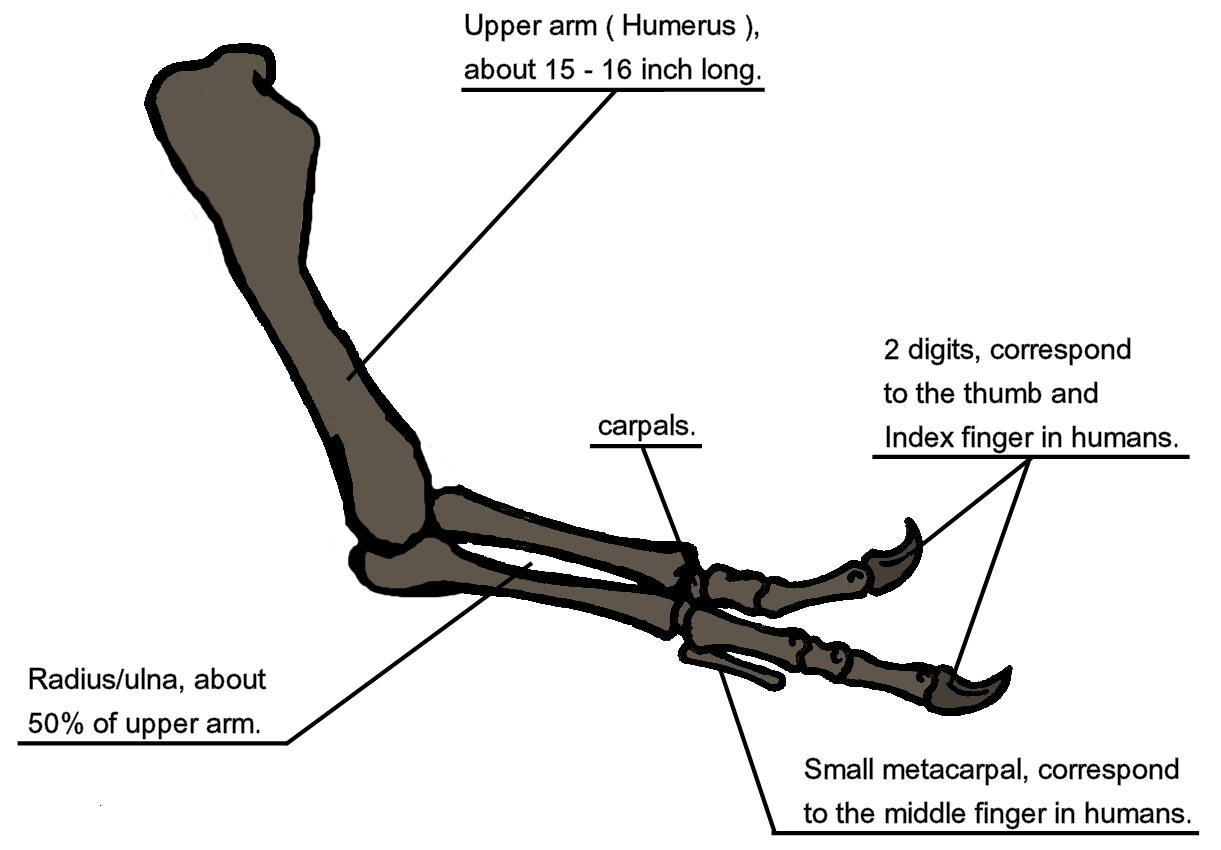
The oldest and most primitive tyrannosauroids are the Proceratosauridae. The oldest known are Middle Jurassic English Proceratosaurus and equally old Siberian Kileskus. Slightly younger (and far more completely known) is Guanlong of the Middle-Late Jurassic boundary of China: a 3 m or longer crested proceratosaurid. Like other early coelurosaurs, the arms were fairly long. Long arms with tridactyl manus were likely found in all proceratosaurids, but we do not yet have the arms for Proceratosaurus, Kileskus, or Early Cretaceous (and possibly 8 m long!) Sinotyrannus, youngest known proceratosaurid.
Dilong of the Early Cretaceous of China represents the next phase of tyrannosauroid evolution. It was the first tyrannosauroid found with protofeathers. At 1.5 m length, it still indicated that some basal tyrannosauroids were small members of the predatory community.
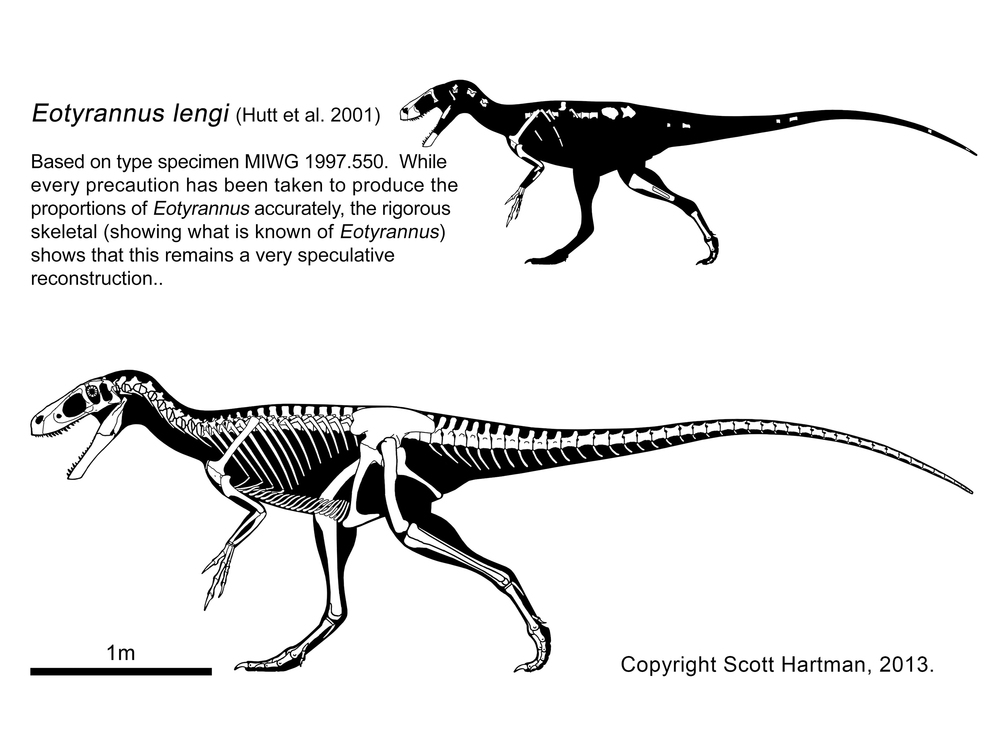
More robustly-built are the "stokesosaurs", including Jurassic tyrannosauroids such as Stokesaurus of North America, and Juratyrant (formerly considered a species of Stokesosaurus) of Europe, and Aviatyrannis of both. A later "stokesosaur" is Early Cretaceous Eotyrannus of Europe, with an adult size of possibly 4.5 m or more. It was dwarfed by other theropods in its community: the carnosaur Neovenator and the spinosaurid Baryonyx.
Newly discovered is Yutyrannus of Early Cretaceous of China. Known from three nearly-complete fossils, this is a 9 m long predator. It still retained the tridactyl manus with long claws of typical tetanurines. Because they were preserved in fine-grained ash-based sediments, the remains of long protofeathers were found over its body, indicating that even giant theropods had a fuzzy coat!
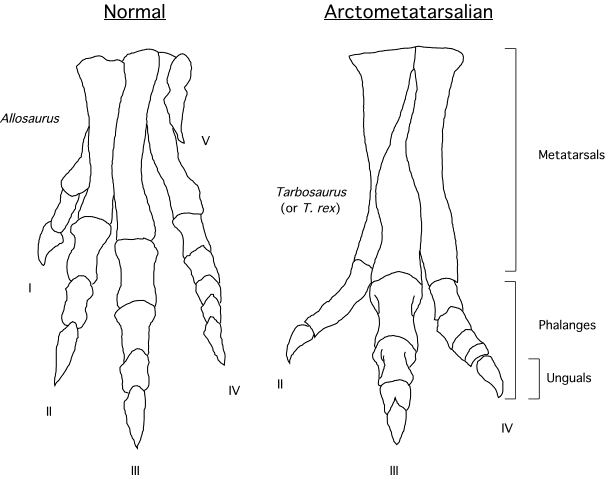
Even larger is 6 m or more longer Dryptosaurus, a late-suriving primitive tyrannosauroid of eastern North America. The arms of Eotyrannus are primitively long; in Dryptosaurus the arm is very short but has a very large claw. It is confirmed as having an arctometatarsus. As with more derived tyrannosauroids, the distal hindlimbs (tibia, metatarsi) are elongated: an indication of cursorial (running) ability. Similar but older is Xiongguanlong of late Early Cretaceous China.
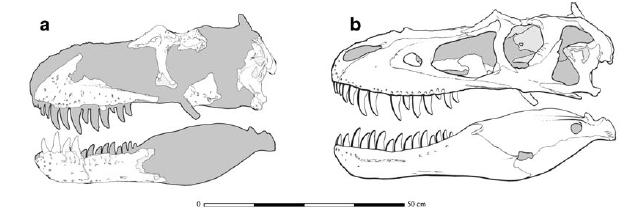
Tyrannosauroids increase size again with the 6 m or longer Appalachiosaurus of the Late Cretaceous of eastern North America and deep-skulled Bistahieversor of the Late Cretaceous of the American Southwest, and again with the Tyrannosauridae proper.
TYRANNOSAURIDAE
Tyrannosaurids proper are one of the last groups of large bodied theropods to evolve, showing up only in the last 20 million
years or so of the Late Cretaceous of North America and Asia. (Consequently they have a similar range distribution to
coronosaur ceratopsians, pachycephalosaurs, corythosaur-line lambeosaurines, and club-tailed ankylosaurids). Although for most of their history
tyrannosauroids were minor predators in their habitats, tyrannosaurids were by far the largest flesh-eaters in their environments.
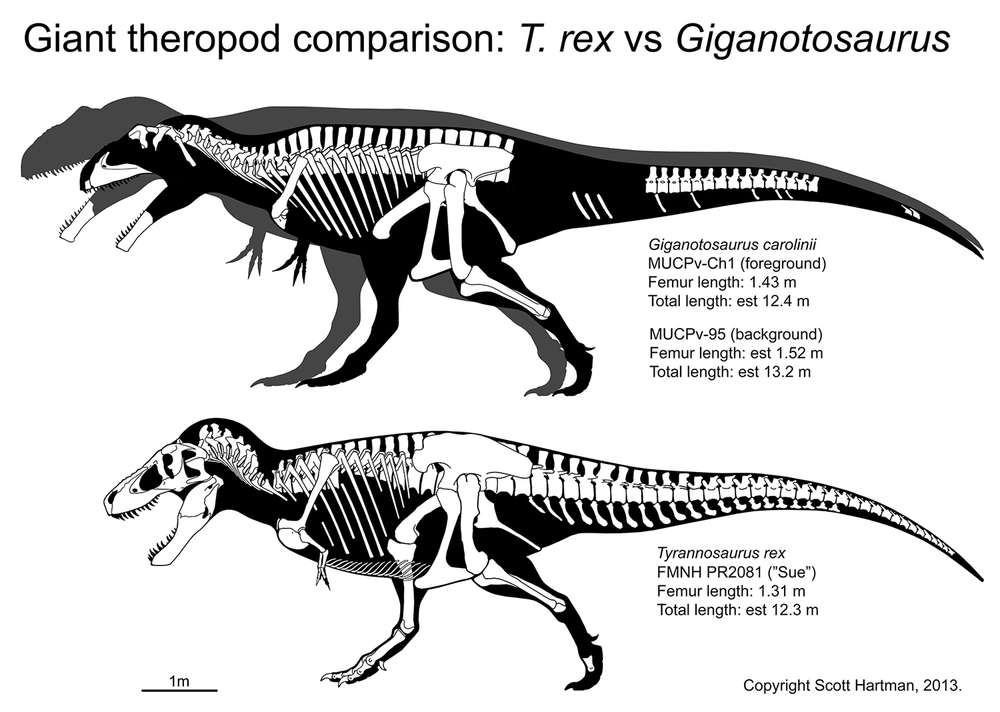
Small tyrannosaurids were about 8 m long; most reached at least 10 m; and at least one genus reached 13 m.
Tyrannosaurids were specialized relative to their ancestors by possessing:
Tyrannosaurids include the relatively slender Albertosaurus and Gorgosaurus of western North America; slender Alioramus of Asia; and more heavily built Daspletosaurus and Teratophoneus of western North America and Tarbosaurus and Zhuchengtyrannus of Asia; and giant 13 m long, 8-10 ton Tyrannosaurus of western North America. Tyrannosaurus rivals the biggest carcharodontosaurs and spinosaurids in mass.
Tyrannosaurids seem to have relied solely on their jaws to kill their food. Their long legs meant that they were faster than their potential prey (hadrosaurids, ceratopsids), although adults of the 2 ton or greater size range may not have been fast runners. (Juvenile tyrannosaurids, though, would have been among the fastest dinosaurs). At least some tyrannosaurids have been found in groups of different ages: possibly family associations.
PLANT-EATING "CARNIVOROUS" DINOSAURS
The remaining coelurosaurs (Maniraptoriformes) all have brains that are twice again as large or larger (based on skull
size) as the more basal coelurosaurs. They also share a suite of unusual features that strongly suggest a move away from the
strictly carnivorous diet of their ancestors and relatives. In particular, they typically:
A new discovery of broad pennaceous feathers (i.e., feathers with a shaft, branches off of that, subbranches off of that, etc.) in ornithomimosaurs seem to place this trait at the same part of the tree as this shift to non-carnivory. However, at present they are only known on the arms of ornithomimosaurs: the spread to other parts of the body seems to be further up the tree.
ORNITHOMIMOSAURIA
The basalmost lineage of the maniraptoriforms are the Ornithomimosauria, the ostrich dinosaurs. Ornithomimosaurs
differ from the ancestral state by:
Their adaptations suggest a move away from predation towards a more omnivorous or even herbivorous lifestyle.
Primitive ornithomimosaurs are known from the Early Cretaceous of Africa (Nqwebasaurus: currently the oldest and most primitive known ornithomimosaur, and the only one from the Southern Continents), Europe (Pelecanimimus), and Asia (Harpymimus, Shenzhousaurus, Hexing, large Beishanlong) and the Late Cretaceous of Asia (Garudimimus, Sinornithomimus).
The latter two share with the Late Cretaceous derived group Ornithomimidae skulls with toothless beaks, and Sinornithomimus shares with Ornithomimidae an arctometatarsus (convergently evolved with Tyrannosauridae): Early Cretaceous Kinnareemimus of Thailand shows an incipient arctometatarsus form. These dinosaurs were among the most cursorial of all theropods.
Ornithomimidae proper includes Late Cretaceous western North American (Struthiomimus, Ornithomimus) and Asian ( Gallimimus, Anserimimus) taxa. Giant Deinocheirus, known only from its arms and a few isolated bones, may be a Tyrannosaurus-sized ornithomimid (or more primitive ornithomimosaur).
At least some ornithomimosaurs lived in herds/flocks. A recent discovery reveals that adult ornithomimosaurs had pennaceous feathers on the arms, but juveniles do not.
MANIRAPTORA
The remaining theropods form the clade Maniraptora ("hand grabbers"). Maniraptorans show numerous specializations:
One possible problematic shared derived feature of Maniraptora is a backwards-pointing pubis. Most coelurosaurs (and saurischians in generally) have a vertically-oriented or anteriorly-oriented pubis. In therizinosauroids, alvarezsaurids, the basal troodontid Sinovenator, dromaeosaurids, Archaeopteryx, and avialians the pubis points backwards; in the basal therizinosaur Falcarius, the basal alvarezsauroid Haplocheirus, oviraptorosaurs, and troodontids other than Sinovenator it points vertically or anteriorly. So it is difficult to say which condition is found in the concestor of Maniraptora. (Regardless of the answer, there is a LOT of convergence going on!!)
Changes in the muscle attachments in the hindlimbs of maniraptorans show a switch from the femur-and-tail power stroke found in other dinosaurs (inherited from the early diapsids) to one where the flexion of the knee is more important.
Maniraptorans are the most diverse clade of dinosaurs. None retain a basal theropod form: indeed, very few retain the ancestral carnivorous condition. Major groups include the Therizinosauria, Alvarezsauria, Oviraptorosauria, and Eumaniraptora (which get their own lecture).
The oldest maniraptorans are some possibly Middle Jurassic eumaniraptorans, and definitely this clade is present by the Late Jurassic. A therizinosaur dentary possibly from the Early Jurassic (but may be as young as the Early Cretaceous!) of China is considered by some authors to be a therizinosaur: however, it might simply be a derived sauropodomorph.
THERIZINOSAURIA
This group and the Oviraptorosauria were once thought to form their own clade (Oviraptoriformes). However, discovery of the primitive
members of both Therizinosauria and Oviraptorosauria shows that many of the similarities between the derived members of these clades
are convergences. More recent studies typically place therizinosaurs as the most basal branch of Maniraptora.
Therizinosauria ("scythe reptiles") have been considered sauropodomorphs and late surviving proto-ornithischians, but are in fact coelurosaurian theropods. Prior to the 1990s, they were often called the "segnosaurs." Similar to the ornithomimosaurs, this group is characterized by:
Other than the jaw Eshanosaurus, this group is known only from the Cretaceous, and only from Asia and North America at present. The basalmost form is Early Cretaceous Falcarius of western North America. It retains a relatively elongate metatarsus and a vertically-oriented pubis. The derived therizinosaurs form the clade Therizinosauroidea, and are characterized by shortened metatarsi in which all four toes touch the ground and backwards-pointing pubes. (In this case, like the ornithischians, this is almost certainly to accomodate a large gut for digesting plants.
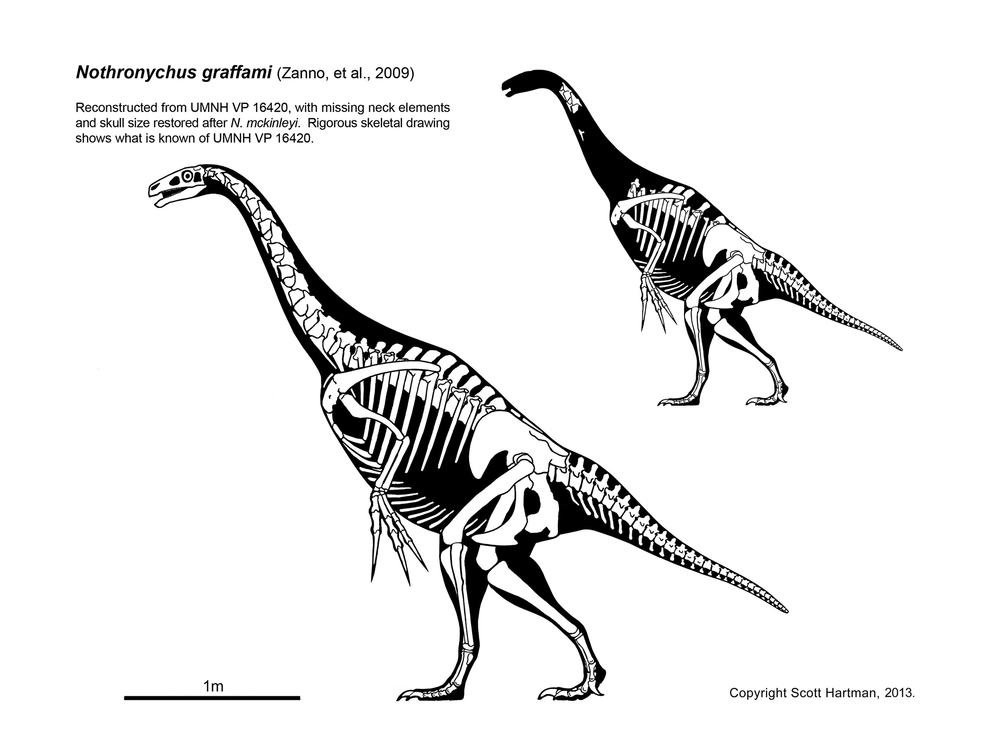
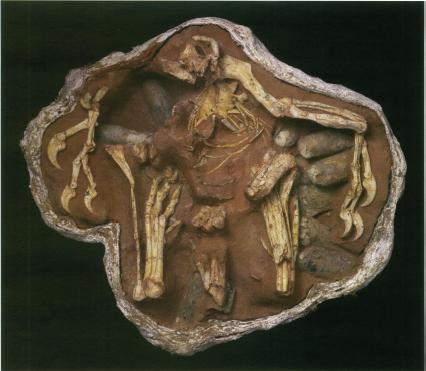
Therizinosaurs seem to have been primarily, if not strictly, herbivores. Their stumpy feet and short legs show them to have been among the slowest theropods. To defend themselves (and possibly to help them feed) they had huge claws. They ranged from bear-sized taxa such as Erlikosaurus and Beipiaosaurus through Nothronychus to Tyrannosaurus-sized Therizinosaurus with 1 m long claws.
ALVAREZSAURA
Alvarezsauria (sometimes called "Alvarezsauroidea") is a recently discovered, highly specialized group of maniraptoran theropods.
Haplocheirus is the oldest known form, from the early part of the Late Jurassic of China; otherwise, the remaining alvarezsauroids
(collectively the Alvarezsauridae) are from the Late Cretaceous. Alvarezsaurids are known from
South and North America, Europe, and Asia. They have numerous bird-like features, and were once thought to have been
specialized flightless birds. Alvarezsaurids range in the chicken-to-rhea sizes.
Alvarezsaurids have small beaky skulls with tiny teeth and hands in which the thumb is much more powerful than the other fingers.
In the Cretaceous Alvarezsauridae, the forelimbs are further transformed into bizarrely poweful arms with a huge thumb claw and exceedingly small digits II and III. The alvarezsaurids have a backwards pointing pubis. Unlike the therizinosauroid and ornithischian situation, this backwards position of the pubis is more likely associated with changes in the locomotory muscles towards knee-driven power from the ancestral tail-and-femur driven power.
Only a little is known of Alvarezsaurus itself (the basalmost form); somewhat more is known for the more derived Patagonykus and Achillesaurus (all from South America). Only a foot is known of rhea-sized Kol of Asia.
The highly derived Parvicursorinae (also called "Mononykinae"), in contrast, are known from many excellent specimens. The best studied are the Asian taxa Mononykus, Parvicursor, and Shuvuuia. More fragmentary Asian parvicursorines include tiny Albinykus, Linhenykus (in which digits II and III had entirely vanished), and Xixianykus. (However, North American forms such as Albertonykus are known). Parvicursorines have an extreme version of the arctometatarsus, in which the upper portion of metatarsal III is entirely missing.
The parvicursorines show numerous cursorial adaptations, but these were almost certainly defensive. They seem to have been insectivores, and their forelimbs may have been used to batter into ant and termite nests. They have been found from deserts to well-watered environments.
The remaining maniraptorans are the oviraptorosaurs and the eumaniraptorans. These two groups are united by several important characteristics:
OVIRAPTOROSAURIA
Another odd batch of maniraptorans is the Oviraptorosauria. This group is characterized by
The basalmost oviraptorosaurs are toothy Incisivosaurus, Protarchaeopteryx, Similicaudipteryx, and Caudipteryx, all from the Early Cretaceous of China.
Other early branches of the oviraptorosaurs are Microvenator of the Early Cretaceous of western North America and specialized Avimimus (with an arctometatarsus) of the Late Cretaceous of Asia. (The latter seems to have dwelt in herds/flocks/whatever).
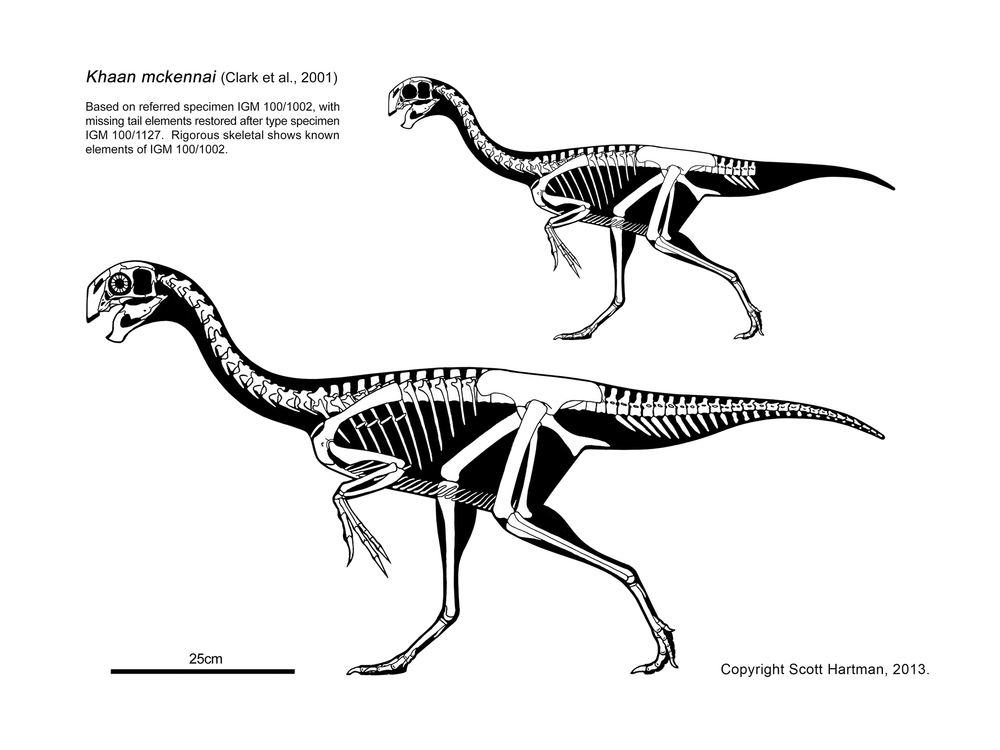
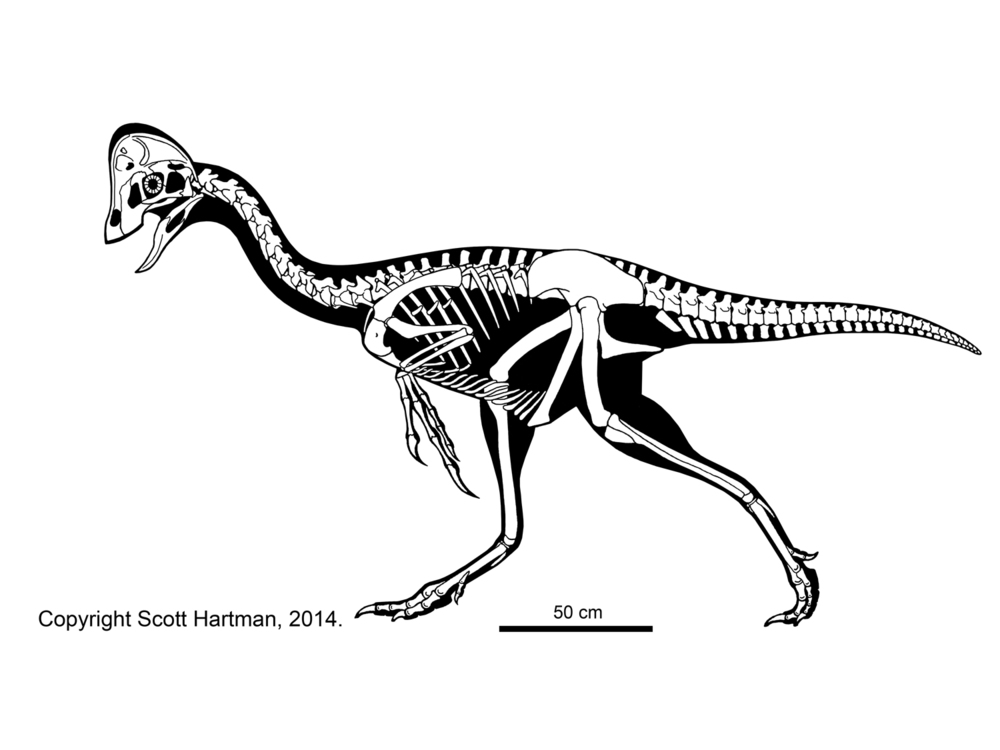
These basal branches of Oviraptorosauria are relatively small (chicken-to-turkey sized). The more derived Caenagnathoidea contain forms that range from turkey to human to tyrannosaur size. Caenagnathoids are derived by loss of all teeth as well as other specializations. Most (but not all) phylogenetic studies divide the Late Cretaceous caenagnathoids into two branches: the Elmisauridae (sometimes called the "Caenagnathidae" or "Elmisaurinae": taxonomy on this remains in flux) which had elongate hindlimbs and sometimes even an arctometatarsus, and the stout-footed Oviraptoridae. However, other studies mix and match the components of these groups. Oviraptoridae proper seems to be limited to Asia, while Elmisauridae is found in both Asia and North America. Examples of oviraptorids include Oviraptor, crested Citipati and Rinchenia, and crestless Khaan. Elmisaurids include North American arctometatarus-footed Chirostenotes, tiny trans-continental Elmisaurus, and a larger not-yet named form from the end of the Cretaceous. By far the largest oviraptorosaur, though, is the recently discovered (?elmisaurid) Gigantoraptor of Asia: as large as an Albertosaurus or other smaller tyrannosaurid.
The life habits of oviraptorosaurs are confusing. While the ancestral ones seem to be convincingly herbivorous, there are lizards in the gut contents of some oviraptorids: perhaps they were omnivorous? Many oviraptorosaurs have been found in desert environments, but others in forested regions.
EVOLUTIONARY PATTERNS IN BASAL COELUROSAURS
Feeding adaptation transformations:
Locomotory adaptations:
Gigantism and Miniaturization:
Niche partitioning:
Basal coelurosaurs represented the minor predators of many Jurassic and Early Cretaceous environments. Diversification
into non-predatory modes allowed coelurosaurs to diversify into niches previously unoccupied by theropods. (In
Late Cretaceous Asia, small non-predatory coelurosaurs are very common, while small ornithopods are absent.) In contrast,
tyrannosauroids evolve into giant top predators in the Late Cretaceous of Asia and
North America after the disappearance of carcharodontosaurids and spinosaurids.
To Next Lecture.
To Previous Lecture.
To Syllabus.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
(1).jpg){kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}