
Living dinosaurs (birds) and their closest living relatives (crocodilians) share many derived features of reproduction; these are probably synapomorphies of Archosauria and so would be expected to be found in all extinct dinosaurs:
Fossil dinosaur eggs and nests found in the rock record, as have embryos of most major dinosaur clades.
ALL non-avian dinosaur eggs are basketball-sized or smaller: NO dinosaur hatched from eggs the size of people!!
Some dinosaur nests associated with covered mats of vegetation: probably helped to keep warm (as in croc nests).
Some dinosaurs (maniraptorans) found in "brooding position" over nests; unlikely to be found in dinosaurs which are too large (i.e., tyrannosaurids, hadrosaurids, sauropods, etc.) or lacking feathers (non-coelurosaurs). An ongoing study of the density of pores on archosaurian eggshells shows that most dinosaurs have characteristics of eggs from covered nests, but that maniraptorans show open nests. This is strongly consistent with the record of brooding.
In primitive modern birds it is the male rather than the female which broods the nest: paternal care. These nests are laid by multiple females. In these paternal care cases, the male rather than the female typically watches over the young after they hatch. When plotted against body size, the volume of eggs of nests of deinonychosaurs and oviraptorosaurs more closely resembles the pattern seen in paternal caring birds than in maternal caring or biparental caring birds; this suggests that paternal care may be an ancestral (maniraptoran?) trait that was passed on to primitive avians but modified in more derived birds. However, we do not have evidence yet to see if more basal dinosaurs had paternal care, maternal care (as in crocodilians), or biparental care. And indeed the statistics supporting the paternal care hypothesis in non-avian maniraptorans has been called into question.
Most primitive modern birds are ground nesters; suggests that tree nesting did not evolve until well into the modern bird (Aves) radiation.
Dinosaurs tend to have nests of about a dozen or so eggs each: more than found in modern birds, less than in (for example) turtles. This is regardless of size: troodontids to titanosaurids!
Implies that unlike placental mammals, dinosaurs could produce a dozen or so offspring a year regardless of size; among placental mammals, larger body size means LONGER gestation periods.
Two main potential life habits upon hatching:
Some evidence of these habits in hatchling dinosaurs:
Parental care of babies for at least several weeks is present in both modern crocodilians and modern birds, implying that this trait was present in archosaurs ancestrally. Birds have long been known to provision (bring food to) their babies; there are cases of crocodilians in captivity doing the same (see video below), but it is not yet known if they do this in the wild. But certainly crocodilian mothers bring their babies to safer places in ponds and lakes where they little ones could hunt.
After baby dinosaurs left the nest, at least seem to have travelled together. Packs/herds of of baby ankylosaurids, ceratopsians, and ornithomimosaurs are known; in at least the case of the ceratopsian Psittacosaurus and the ornithomimosaur Sinornithomimus these herds can include individuals of different ages, including adults. ( The parents may have been with the other baby herds, but were too big to be buried.)
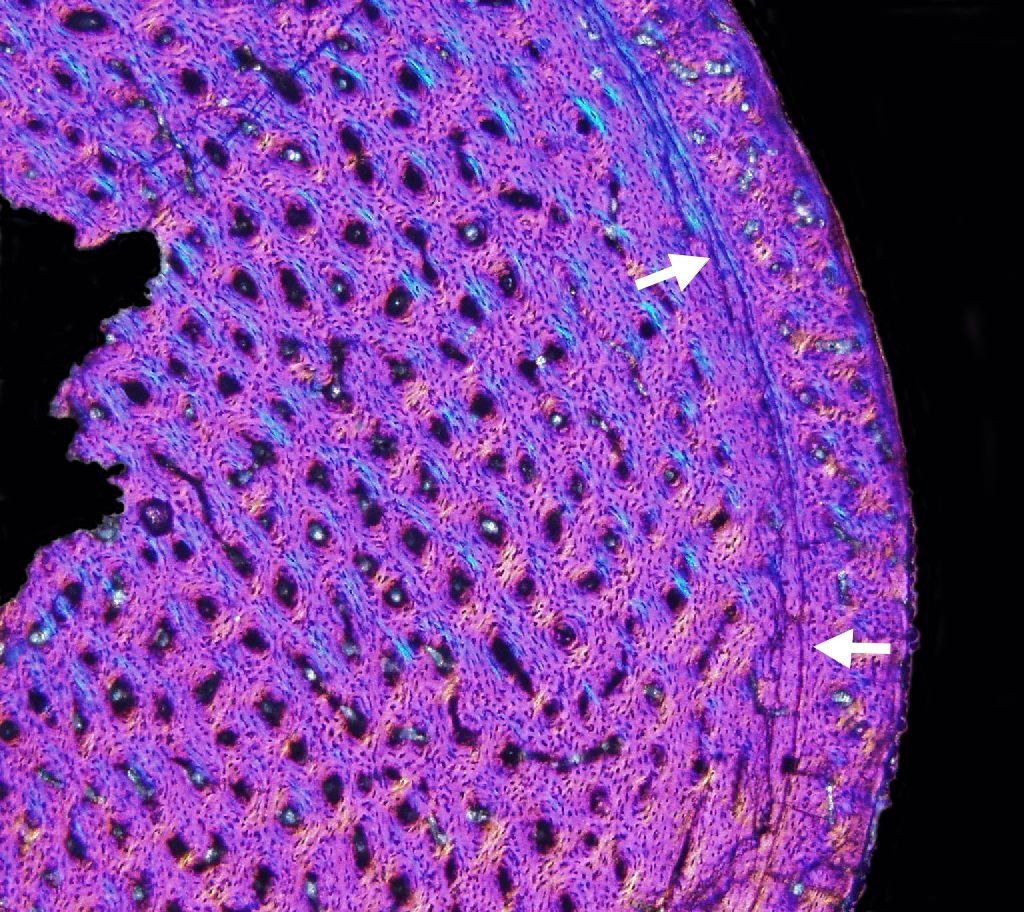
Dinosaur Growth Rate and Skeletochronology
Like many animals, dinosaurs deposited Lines of Arrested Growth
(LAGs for short). These are basically growth rings like tree trunks, one per year. Various paleontologists use that information to
"age" individual dinosaur fossils. By comparing the age of the dinosaur to its size, and plotting all individuals of the species on a graph,
you can figure out the growth pattern and growth rate of that dinosaur species: that is, we can develop a skeletochronology for some species.
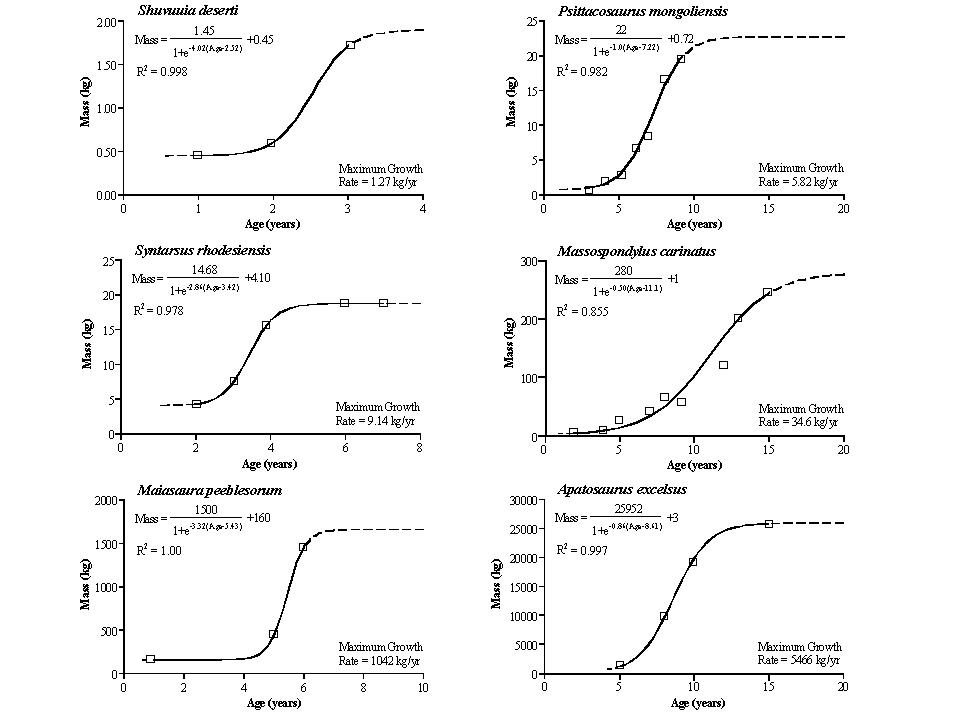
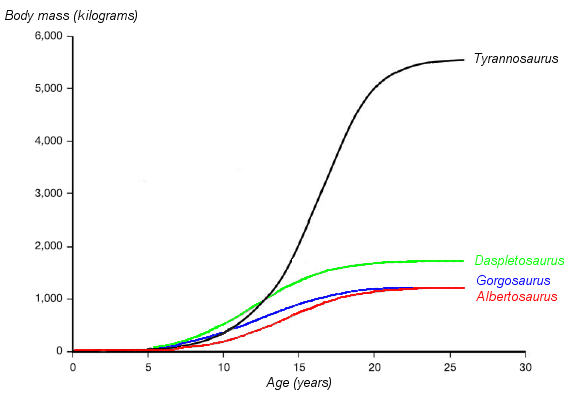
Actual rate was VERY HIGH compared to typical reptiles, particular in big dinosaurs. Estimates based on bone "growth rings" (and other features) indicate small dinosaurs were full grown at only a couple of years, hadrosaurids (e.g. Maiasaura) at only 7 years for, and only 15-20 years for big sauropods (e.g., Apatosaurus) and theropods (e.g., Tyrannosaurus) to reach adult size.
(In contrast, big crocodilians from the Late Cretaceous seem to have taken 50 years or so to reach the same size as big hadrosaurs).
Since most animal populations stay generally stable over time, more baby dinosaurs died before reaching adult size than in typical populations of modern birds or mammals (imagine herd of antelope where every female produced a litter of 12 every year!).
Unlike typical non-avian reptiles, dinosaurs seem to have determinate growth: reaching a fully adult size, than stopping growth (or at least slowing it WAY down: all the LAGs bunch up together).
Lifespans for dinosaurs seem to be shorter than those of similar sized mammals. For instance, the oldest known Tyrannosaurus individual is only 28, the oldest known sauropod studied so far is only 38, but elephants can live to 70 or more.
Life-History Strategies
Different types of organisms have different life history strategies: different ways of dealing with how many young are born at a time;
how many survive to adulthood; how long they live; etc. There are two general types of strategies that we see commonly in nature:
Non-avian dinosaurs are definitely more r-selected than typical larger placental mammals, but more K-selected than sea turtles.
There are lots of unsupported, less-than-reasonable, pseudoscientific explanations:
So, let's take a look at other possibilities.
Was It Limb Posture (i.e., the Parasaggital Stance)? It is true that all the supergiant land animals (dinosaurs, elephants, deinotheres, indricothere rhinos, etc.) all have an upright stance. While there are giant 10-15 t crocodilians contemporaneous with and after the Age of Dinosaurs, these semi-upright animals were primarily aquatic and in a sense don't count.
But other groups with parasaggital stance--such as most crurotarsans, marsupial mammals, modern birds, etc.--never reach these sizes. So it might be that a parasaggital stance is necessary but not sufficient for the >10 t size.
Was It From a REEAALLLLYYYYY LOOOOONNNNNGGGGG Period of Growth? Giant tortoises and big modern crocodilians live a very long time (centuries in the case of tortoises), growing throughout their life. Perhaps dinosaurs just grew for a tremendously long time? But as we saw previously, dinosaurs had a rapid growth rate and relatively short life spans.
So we can REJECT the idea that a long growth phase led to giant dinosaur size!
Very Efficient Dinosaur Hearts and Lungs? Yes, they had both, as we will see. These were probably a necessity for giantism, but not sufficient.
Was it Exceedingly Short Food Chains? When we watch nature specials on TV that focus on large charismatic modern land mammals, we get the impression that the simple food chain of plant → large herbivore → large carnivore (e.g., grass → zebra → lion) is typical. In fact, while it does apply to the world of Cenozoic hoofed mammals and their predators, it actually is not the case for most terrestrial life. As in the marine realm, most terrestrial food chains have many steps from primary productivity to apex carnivore, often through insects and other herbivorous invertebrates, to carnivorous arthropods, to small-bodied insectivorous animals, to predators of those, and so on. (For example, plants → aphids → ants → spiders or mantids → small insectivorous birds → snakes → hawks). And of course there is loss of energy every step up the food pyramid.
Ecosystems of protomammal synapsids of the Carboniferous, Permian, and Triassic, and perhaps even protocroc crurotarsans of the Triassic, seem to have had relatively few large bodied herbivores, and may have had these longer, energy-leaking food chains. And thus, there is less energy available towards the tops. In contrast, dinosaurs produced a broad variety of herbivores able to capture the energy at the base of the pyramid, with a set of carnivores feeding on these. These short food chains may have allowed much more of the solar energy to make its way into meat.
Of course, there is still the observation that even mega-mammals with similarly short food chains fail to achieve dinosaurian size, so this cannot be the entire story. However, it is worth noting that said mega-mammals are the ONLY other case of terrestrial vertebrates which get anywhere near dinosaurian size.
Was it Changing Atmospheres? Even as dinosaurs were evolving, the atmosphere they were breathing was evolving, too. Geochemists have seen that the ratios of various gases, including oxygen, have varied over geologic time. At least some models suggest that the Middle Jurassic though the end of the Cretaceous had oxygen levels exceeding the present 20%. This would mean that every breath a dinosaur took would have more oxygen, making it easier to power a high metabolism.
Furthermore, experiments of growing plants of Mesozoic varieties under Mesozoic-style atmospheres suggests that their productivity (essentially, the amount of nutrients they produce per area per unit time) could go up 2 to 3 times present day conditions. If so, then there would have been more food available per unit area for the herbivores (and from this up the energy pyramid), again making it easier to be an endotherm in these conditions.
Environmental changes in Mesozoic discussed above may have supported giantism:
The second aspect is probably easier to deal with. There are several possible advantages:
However, we may be asking the wrong question. The problem might not be "how did dinosaurs get so big?"; it might be "why DON'T placental mammals get so big?" This may in part be due to the lower oxygen level and productivity during the Cenozoic. However, it may have to do with mammalian vs. dinosaurian reproduction:
In contrast, even giant dinosaurs could could lay clutches of a couple dozen eggs a year or more. Thus, they could survive environmental change much more easily than big mammals. So mammals may not be able to reach giant size because they become extremely vulnerable to extinction.
Additionally, the thicker cartilage of dinosaurs was present at all body sizes, whereas the size of the articular cartilage in mammals decreases with increasing body size. As a result, larger mammals have less cushioned joints, which may have been a hinderance to activity. So it may simply be that larger land mammals could not actually move effectively, but that dinosaurs continued to be mobile.
To Next Lecture.
To Previous Lecture.
To Syllabus.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}