The Early Cretaceous Asian dromaeosaurid Microraptor and the avialian Jeholornis by Apsaravis
Key Points:
•While gliding is very common among living things, powered flight is rare and only evolved in insects, pterosaurs, birds, and bats.
•Unlike bats and pterosaurs (and scansoriopterygids), bird and their outgroup did not primarily rely on skin for their flight surface, but feathers.
•Up through the beginning of the 21st Century, two rival models were proposed for the origins of flight: either from the trees-down of from the ground up.
•Discovery that modern birds use a series of wing-based non-flight behaviors (such as wing-assisted incline running (WAIR) and controlled-flapping descent (CFD) suggests that the trees-down vs. ground-up dichotomy is false.
•At least some of the non-avialian eumaniraptorans seem to have been capable of powered flight; however, many early eumaniraptorans (including early avialians) probably only had a limited capacity.
Last lecture we saw the origin and evolution of flying dinosaurs ("birds" in the broad sense), and before that the origin of feathers.
But we haven't yet addressed the origin of flight, nor how all these things are related to each other.
Evolution doesn't work by massive instantaneous transformations: a fully flying modern style bird did not hatch out of a totally non-flying
dinosaurs egg! Instead, each phase along the way functioned in its own way, well enough for that dinosaur to make a living at it. And (as
we'll see) it wasn't as if dinosaurs were trying to become birds; instead, each adaptive phase produced several divergent branches,
some evolving in one direction and others in others.
Our knowledge of the phases of the origin of bird flight (anatomical, phylogenetic, and behavioral) have all greatly increased in the
last decade. But before we see this newer understanding, let's review some terms and take an historical look at the problem.
BACKGROUND:
Some key terms:
Arboreal: tree-dwelling (spends most of the time in the tree branches)
Cursorial: running
Integument: body covering, such as scales, feathers, fur, etc.
Scansorial: tree-climbing (spends most of the time on the ground, but will climb up into trees to rest, hide, etc.)
Powered flight: capable of generating lift and thrust with wings in order as it travels through the air (as opposed to gliding or
parachuting, which might start with a powered leap but only use gravity and air currents once they are in the air)
Volant: flying
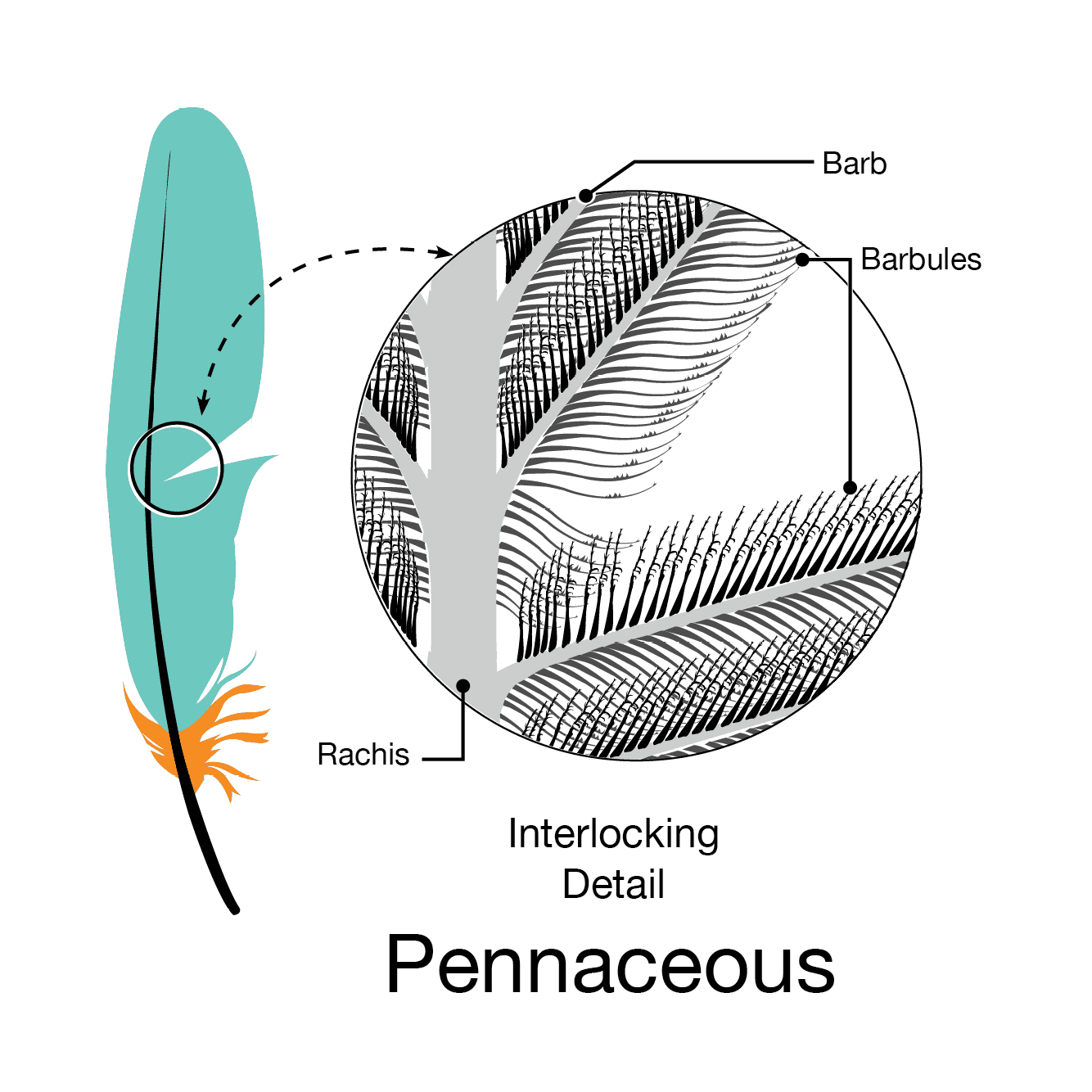
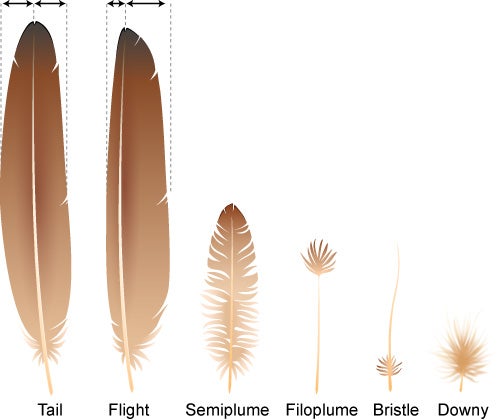
Pennaceous: classic feather form, with a vane down the middle. Includes flight feathers, contour feathers, and some display feathers
Plumulose: feathers lacking a major vane. Includes down (the typical covering of modern baby birds) and the "protofeathers" of non-maniraptorn dinosaurs.
In the history of life, only four groups of animals have evolved powered flight:
Insects: the first fliers, from the Carboniferous Period onward
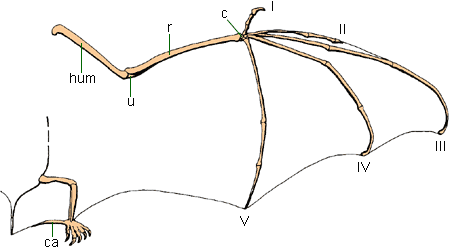
Pterosaurs: the first vertebrate fliers, from the Late Triassic until the end of theCretaceous. The wing is formed by a membrane stretched between the elongate arm (with an extraordinarily long digit IV), the hindlimbs, and the body
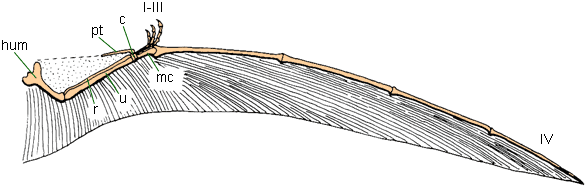
Birds: if we mean "fully powered flying birds", then that is probably Ornithothoraces, and thus from the Early Cretaceous onward. Bird wings are formed by feathers attached along the elongate forelimb
Bats: from the Paleogene Period onward (oldest bats about 55 Ma). Bat wings are formed by skin stretched between elongate arms with especially elongate digits III-V, the hindlimb, the tail, and the body.
Traditionally, paleontologists have considered two main scenarios for the origin of bird flight:
"Trees Down" (also called the Arboreal Model)
Ancestors of birds were tree-dwellers (arboreal)
Powered flight evolved from gliding/parachuting:
Tree-dwelling animals jumped from branch to branch
Those with a gliding/parachuting surface could travel further, so selection favored development of increased wings
Eventually, forms modified the gliding surface to give them additional thrust: powered flight
Seems like a reasonable scenario for the origins of the other powered flying vertebrates (pterosaurs, bats)
Makes sense energetically, since the early gliding phases can use gravity to help them fly long before the need for the development
of strong arm muscles
As seen above, gliders/parachuters are VERY common, and tetrapod gliders/parachuters have consistenly evolved convergently from
arboreal animals
"Ground Up" (also called the Cursorial Model)
Ancestors of birds were ground running animals (cursorial)
Powered flight evolved from activity useful to runners, outside of the context of a tree-dwelling phase
Evolution of the wing stroke evolved in some non-flight context (possibly food capture; possibly as a speed-aid or an aid for
leaping and jumping)
Feathers originally evolved in a non-locomotion context, but were exapted for whatever the possible pre-flight use of the forelimbs
was
Through enlargement of the proto-wing in the non-flight context, the forelimbs became large enough and developed enough to begin
to carry the animal through the air
Birds only got into the trees after having developed the early phases of flight
Birds, unlike bats and pterosaurs, do not make use of a membrane to fly; and there is no fossil evidence that they ever did
Unlike bats and pterosaurs, the hindlimb is not part of the flight surface; in fact, most modern and fossil birds have perfectly
good running legs (just as their outgroups had)
Until recently, all the known member of avialian outgroups (Dromaeosauridae, Troodontidae, Oviraptorosauria, Therizinosauria, Ornithomimosauria) were fairly large bodied animals that were unlikely to have spent much time in trees
But there was a lack of good modern analogues for whatever the ground use scenario would have been
Also, some questioned whether it would have been energetically feasible for animals to achieved powered flight directly from a running/leaping
behavior
(NOTE: during the 1970s-1990s, this debate was tangled up with a scientifically separate debate; that is, where birds fit phylogenetically
among the archosaurs. The media in particular made the equation "'arboreal model = non-dinosaurian origin of birds'; 'cursorial model =
dinosaurian origin of birds'". But these were actually separate debates. Even among those who recognized the dinosaurian origin of birds,
some argued for the trees-down model, and others for the ground-up.)
This debate was primarily waged prior to the new discoveries of Early Cretaceous feathered coelurosaurs, which greatly increased our
knowledge of the anatomy (integumentary and skeletal) of the basal members of the coelurosaur clades. Additionally, important observations
of modern birds revealed a very significant locomotory behavior, previously overlooked.
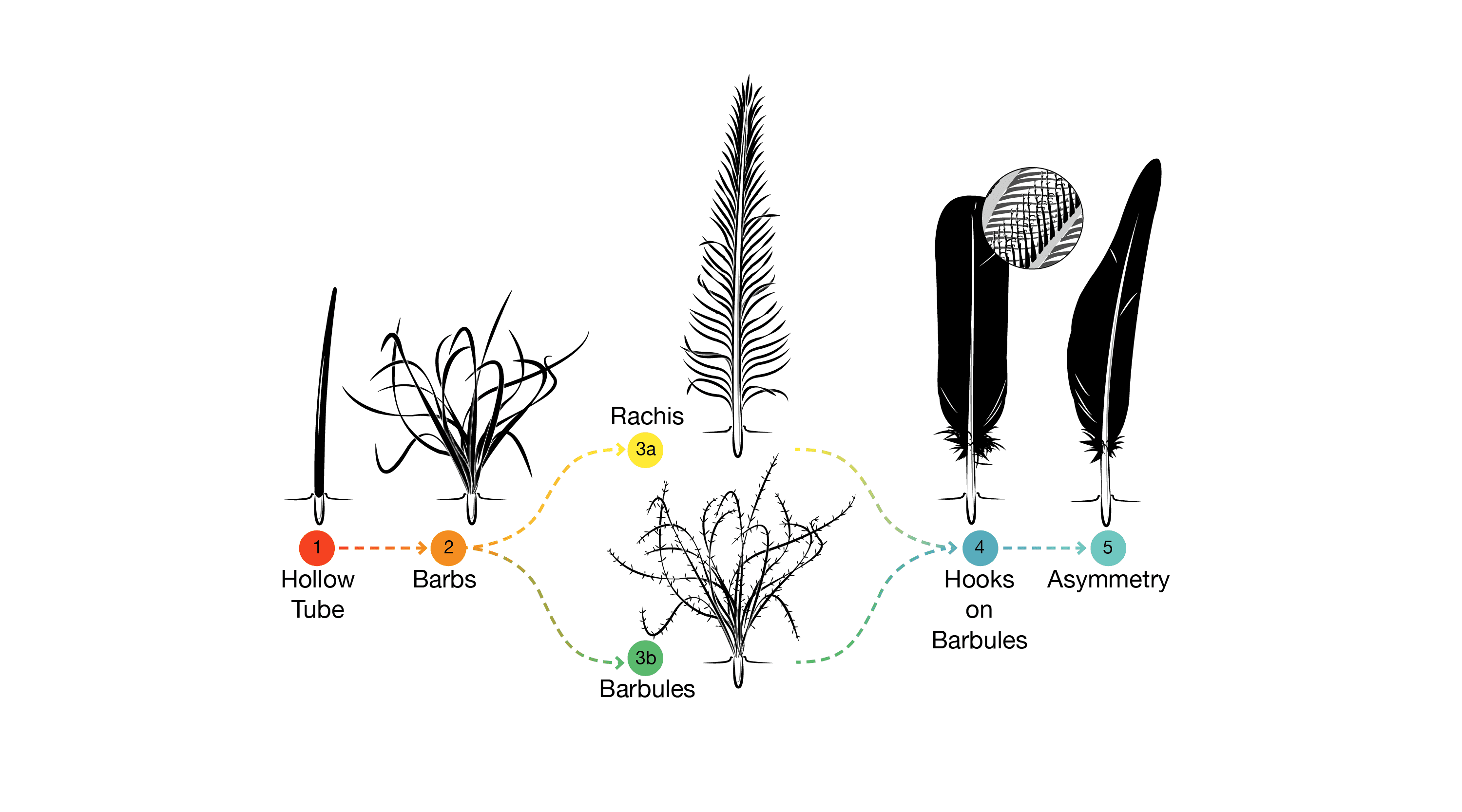
EVOLUTION OF FEATHERS: Modern feathers are very complex structures, with a hollow shaft that branches, with those branches branching and the branches of the branches branching. Modern feathers are NOT just the big flight feathers of the wings and tail: there are body feathers (that insulate and streamline the body); down (that keeps pre-flying baby birds warm); display feathers; and others.
Until recently, the basalmost theropod known to have feathers was Archaeopteryx, although some researchers speculated that other theropods had them as well. And the feathers of Archaeopteryx were identical to the feathers of modern birds, so they didn't reveal much about the early phases of these structures. But fossils from lake sediments of the Early Cretaceous (and now the Middle Jurassic) of China have given us a better understanding of the distribution of feathers and protofeather structures.
Primitive theropods (and now primitive ornithischians) show a body covering of plumulose protofeathers: simple hair-like or down-like filaments. Based on the developmental biology of modern feathers, this is the expected earliest phases of feather evolution. Protofeathers obviously don't have a flight function, since they don't form an aerodynamic surface. However, they might serve other functions:
Insulation (like fur of mammals and body feathers of birds)
Display (particularly as they might have been brightly colored and/or patterned)
Cover for brooding eggs
Or a combination thereof
However, at least some of the protofeathers of primitive theropods (such as the tyrannosauroid Yutyrannus) are long strap-like pennaceous feathers of the sort also found in maniraptorans. The poorly preserved feathers of ornithomimid arms and tails are also pennaceous, apparently broader than in more basal theropods but not as much as in pennaraptorans.
Note that the fuzz of the heterodontosaurid Tianyulong is very similar and some of the structures on Kulindadromeus are as well. (Kulindadromeus had other integumentary structures not yet found in any other dinosaur.) Whether these turn out to be formed by convergence with theropods or retention in both groups of some ancestral dino-fuzz is not yet certain. (If it is a homologous structure, this means that the concestor of all Dinosauria was fuzzy, at least in part!)
Plumulose protofeathers of this sort seem to be present as part of the body covering of therizinosaurs, oviraptorosaurs, dromaeosaurds, troodontids, and basal avialians. (In ornithothoracines, they are replaced by body feathers.)
Tyrannosauroids and therizinosaurs in addition have strap-like protofeathers intermediate in size and form between the downy protofeathers and true feathers. At present it is not certain what the exact structure of the integument of alvarezsaurs was like. However, these pennaceous strap-like feathers (as well as fully modern feathers) are also present in oviraptorosaurs.
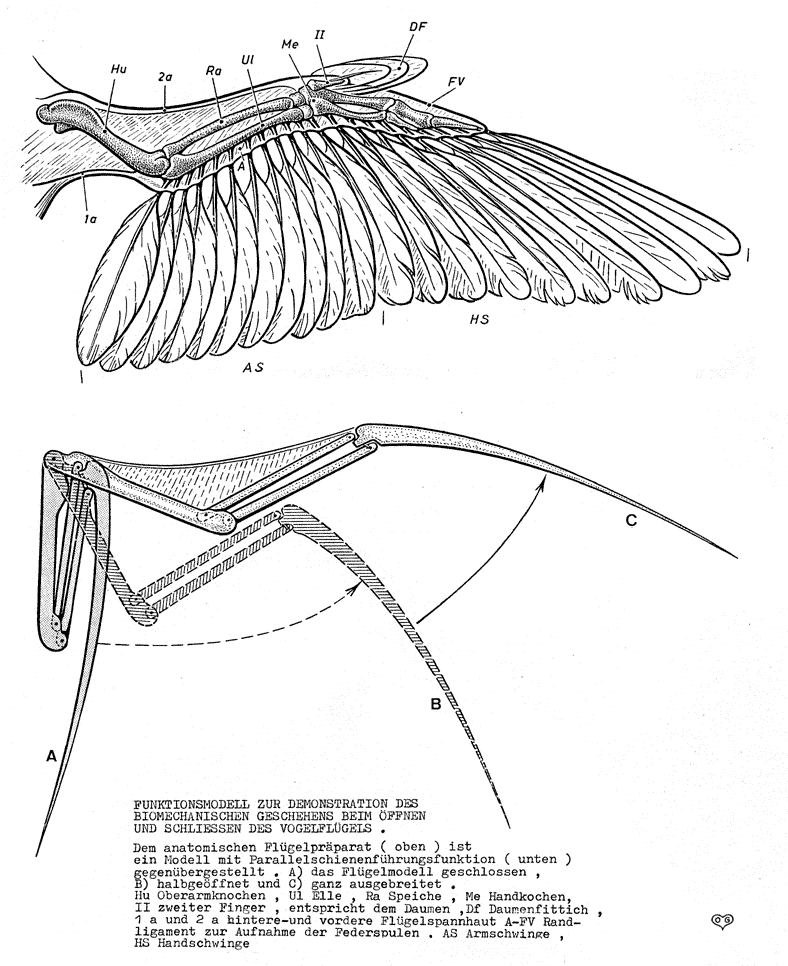
All pennaraptorans for which the integument is preserved (oviraptorosaurs (including scansoriopterygids), dromaeosaurids, troodontids, avialians) show true broad pennaceous feathers on the arms and the tail. In eumaniraptorans (dromaeosaurids, troodontids, and basal avialians) long broad true feathers are present on the hindlimbs as well. Additionally, these same dinosaurs show some major transformations of the forelimb: elongate arms; semilunate carpal allowing for folding the hands; laterally oriented shoulder joints allowing the arms to stick out sideways; enlarged sternum for more powerful arm muscles. In modern birds these adaptations are useful in the flight stroke.
Were all these dinosaurs fliers? It appears unlikely, given the anatomy of the non-eumaniraptoran forms especially! What other function could broad feathers have served?
Maniraptorans are known to have brooded their eggs (nests with parents on them are known for oviraptorosaurs, troodontids, and avialians). Broad feathers may have helped insulate and protect these eggs: increased arm length, better ability to stick arms out to the side; increased surface area of feathers allows greater coverage of eggs. So perhaps brooding was a selective feature in these adaptations?
Additionally, the carnivorous members of this clade used their forelimbs to capture prey: elongation of the forelimbs gave them a better range in which to strike, while modifications of the shoulder and wrist would let them fold the arms in tight rather than drag them along.
However, it is observations of modern birds that revealed another possible selective force in the evolution of the maniraptoran forelimb and feathers.
WING ASSISTED INCLINE RUNNING (WAIR) and CONTROLLED FLAPPING DESCENT (CFD):
In the early 2000s research by Ken Dial of the University of Montana's Flight Laboratory revealed a locomotory behavior in modern birds not previously realized. Birds (in this case chukar partridges) were discovered to
run vertically up surfaces, aided by flapping their wings back and forth in order
to generate traction against the surface. They called this behavior Wing Assisted Incline Running, or WAIR for short. They have since found many bird species with this behavior, even perfectly good fliers like pigeons:
Note that these birds are NOT climbing in the typical sense: they are literally running up the sides of trees. Dial and his team studied the
ontogenetic (growth) changes in the ability for birds to use this behavior, and also experimented by trimming the feathers of birds
to different lengths.
They found:
As the bird wing develops longer, the bird can WAIR up steeper and steeper slopes
As the size of the bird's wing feathers increases, the bird can WAIR up steeper and steeper slopes
The motion used to generate thrust during WAIR is different than that used in flight; in fact, the motions of at least the juvenile form of this locomotion was likely within the scope of typical maniraptorans.
Additional work has shown that this behavior is widespread among modern birds.
The apparatus required to use WAIR is:
Elongate arms capable of a broad front-to-back stroke
Reasonable good chest muscles to power this stroke
Broad feathers on the arms
Hindlimbs capable of running
All these attributes are present in many pennaraptorans (oviraptorosaurs, dromaeosaurids, troodontids, basal avialians). Additionally, modern birds use WAIR to escape predators: certainly a selective factor present in the Mesozoic, too! Furthermore, there are net selective advantages to slight increases in the length and breadth of the feathered arm surface: the sort of material that can easily be increased by natural selection.
WAIR might represent a "stepping stone" or "behavioral missing link" in the origin of flight. Small (or juvenile) maniraptorans might have used this method to escape predators. Now that they had the ability to get up into trees and other high spots, some lineages of maniraptorans might become specialized for life up on these high spots. Additional natural selection could favor further development of wing size and shape as an aid for getting back down off of high places (controlled flapping descent), or (eventually) from branch-to-branch.
Thus, WAIR serves as a functional link between cursorial and arboreal models (and organisms). It is a cursorial model in that wings begin in part as an aid to running locomotion (just vertical running); it is an arboreal model in that once maniraptorans have an ability to get into the trees, evolution can further develop the forelimbs to get them back down to the ground. And all of these behaviors are still found in modern animals: no speculation of behaviors not currently seen needed.
A NEW SCENARIO FOR BIRD FLIGHT ORIGINS:
The various recent discoveries of the skeletal and integumentary anatomy of various coelurosaurs (including basal avialians) and the behavioral and biomechanical evidence of modern birds suggests a more complete possible scenario for bird flight evolution than the historical "ground up" or "trees down" versions. Note that as with all evolutionary scenarios this would be a simplification, but the following is consistent with our current evidence:
Phase I: Protofeathered Cursors (Basal tetanurines (or all dinosaurs?)): These dinosaurs retained the long running legs of theropods ancestrally, and had evolved longer grasping arms, larger brains, and increased agility for capturing prey and/or avoiding being captured themselves. Additionally, these dinosaurs (or their more distant ancestors) evolved plumulose protofeathers (for insulation, display, brooding, etc.)
Compsognathids, tyrannosauroids, ornithomimosaurs, therizinosaurs, and alvarezsaurs represent groups showing no real specializations towards a WAIR-related lifestyle, and it is likely that these branches of the coelurosaur tree diverged prior to that specialization. (Note that if Tianyulong's and Kulindadromeus's fuzz winds up having the same evolutionary origin as coelurosaurian protofeathers, than Phase I moves all the way down to the base of Dinosauria!)
As a Phase IB, the evolution of pennaceous feathers (including strap-like protofeathers) by the base of Coelurosauria. However, this doesn't seem to deal with any major locomotory changes, so it doesn't really qualify as a full new phase. Broad pennaceous feathers may have more to do with brooding and/or display than flight.
Phase II: WAIR and CFD (Pennaraptorans): These dinosaurs developed the long folding forelimbs and powerful chest muscles, perhaps initially for increased predatory reach. However, they also evolved broader feathers (perhaps after that initial forelimb increase): maybe in part for brooding, maybe in part for display, maybe in part for WAIR (at least on the arms, if not the tail). Certainly the equipment for limited WAIR is present at this phase. The modifications of the hindlimb (from a femur-to-tail driven system to a knee-flexion one) also occurs at this phase: is it possibly associated with WAIR?
"Classical" oviraptorosaurs represent animals that might have used WAIR and CFD as juveniles, but certainly not as adults. Additionally, their hindlimbs show no real branch-grasping ability.
Scansoriopterygids do show scansorial adaptations. In the case of Yi and Ambopteryx (at least: maybe other scansoriopterygids did, too) there appears to be gliding. However, this is a failed experiment towards flight, not part of the actual sequence which led to avian flight.
Phase III: Tree-Dwelling Aerialists (Basal eumaniraptorans? Or independently within different eumaniraptoran lineages?): A decrease in size, increase of arm length, increase of the arm and tail feathers, development of the leg feathers, modifications of the tail, and development of possibly tree-perching specializations (sickle claw; hyperextensible pedal digit II; distally-placed pedal digit I) suggest a change in life habits. These dinosaurs may have more routinely used WAIR and CFD, even into adult phases for small bodied taxa. The increased wing surface suggests the possibility for aerial locomotion. While some have argued this aerial locomotion is gliding or parachuting, there is no particular evidence for that. (In fact, some form of primitive powered flight is likely here). Elongate leg feathers may have helped the dinosaurs steer in flight, just as today some raptorial birds use them in a dive to help aim when
diving for prey; similarly, the modified eumaniraptoran tail may have helped as a dynamic stabilizer in flight (as well as on the ground, or while in the trees).
Some basal dromaeosaurids, basal troodontids, and basal avialians (i.e., those outside of Ornithothoraces, such as Anchiornis, Archaeopteryx, Jeholornis, omnivoropterygids, and confuciusornithids) represent this phase.
Larger advanced dromaeosaurids and troodontids would have necessarily been flightless, but as juveniles they might have still been able to use WAIR (and even limited flight?) and CFD.
Note that basal "birds" like Anchiornis, Archaeopteryx, Jeholornis, omniopterygids, and confuciusornithids fit within this phase: there is no compelling evidence that they were any better at flight than were Microraptor, Rahonavis, etc. Indeed the shape of their shoulder joints (and lack of sterna in troodontids, Archaeopteryx, and omniopterygids, at least) may have precluded actually complex flapping flight.
Phase IV: Flapping Flight (Basal ornithothoracines): Primitive ornithothoracines like enantiornthines show specializations such as smaller body size and the alula suggesting increased ability to fly. These were almost certainly powered fliers, but still not of modern bird (i.e., avian) flight ability: they lacked the tail fan; they still had long leg feathers; they had smaller pectoral muscles; and so forth.
Phase V (or maybe Phase IV, Part B): Specialized Tail Steering (Basal euornithines): As above, but development of the tail fan suggests much greater aerial ability and landing sophistication. The loss of the long leg feathers seems to coincide with the evolution of the tail fan, suggesting that as the latter developed, the former was no longer needed. (Modern birds lack long leg feathers, with some exceptions like eagles that need very precise diving ability).
Flightless Patagopteryx would have had ancestors of this phase.
Phase VI: Modern bird flight (Basal carinates): Modern style bird flight with powerful pectoral muscles may have been present only at Carinatae, and thus perhaps first appeared only in the Late Cretaceous.
The flightless members of the hesperornithines would be descendants of this phase




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}