
Physiology and metabolism in the fossil record II: Fossils and scenarios
John Merck

Fundamental truths:
- Tachymetabolic endothermic homeotherms greatly outclass bradymetabolic organisms in their ability to marshall energy for hunting, parental care, and other tasks.
- They require an order of magnitude more food per unit time.
This is not an evolutionary transformation that took place quickly or in one great step.
Lingering questions from last week:
- What connection might exist between rates of energy metabolism and bone tissue, the body's reservoir of phosphate ion?
- What connection might exist between the use of modified respiratory surfaces in the elimination of excess heat, and an animal's ability to get oxygen to its cells?
For a discussion of the first question, we turn it over to Dr. Holtz.
(Holtz presents)
Beyond the histology and chemistry of bone, itself, what evidence of thermal metabolic strategies is presented by fossil skeletons? As we examine a couple of scenarios, look for evidence of the following:
- Specific adaptations to ectothermic heat exchange
- Features that seem nonsensical without elevated energy output
- Features that seem nonsensical without the generation of internal body heat
- Improved ventilation of the lungs
- Thermally insulating integument (fur, feathers, etc.)
- Energy-intensive behaviors like parental care of young, play, etc.
A Synapsid scenario:
Constraining the history of endothermy; Tachymetabolic endothermy is considered a characteristic feature of mammals. How did it arise? Somewhere between the first synapsid and the first mammal, the transition from ectothermy to endothermy occurred. Some speculation:
- Dry skin: Frogs and salamanders have little ability to elevate their body temperatures above ambient levels, even by ectothermic means, because of their need to keep their skin moist to facilitate cutaneous breathing. This subjects them to evaporative cooling any time they come into open air. In contrast, even "cold blooded" amniotes can maintain an elevated body temperature while active because their skin is dry and impermeable. Achieving this was the first step in terrestrial thermoregulation. When did it occur? Once indication:
Ectothermic herbivores must maintain high body temperatures to facilitate digestion.
The presence of large herbivorous diadectomorphs and basal synapsids suggests that the last common ancestor of diadectomorphs and amniotes had progressed to the point of elevating their body temperature above ambient levels by ectothermic means.
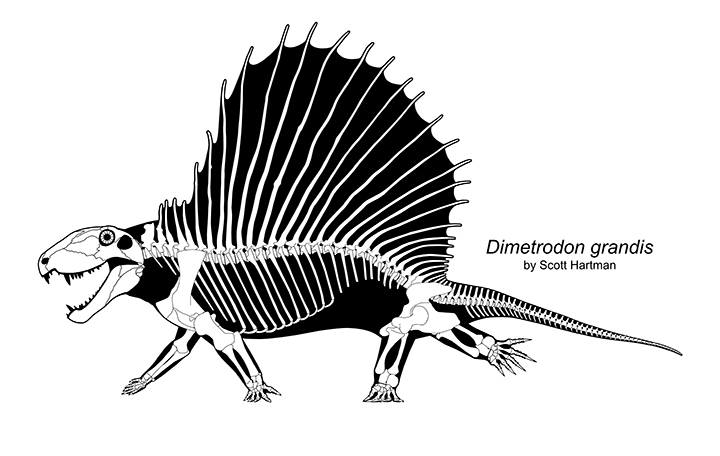
- Cooling fins: Larger animals with lower surface area/volume ratios heat up and cool down too slowly. To accelerate morning activity and remain active at midday, heat exchange surfaces evolved, including the "sails" of sphenacodontines like Dimetrodon. (Of course, this structure could be used for display purposes, also.) But note: The presence of the sail indicates that Dimetrodon lacked the means to either heat or cool itself internally.
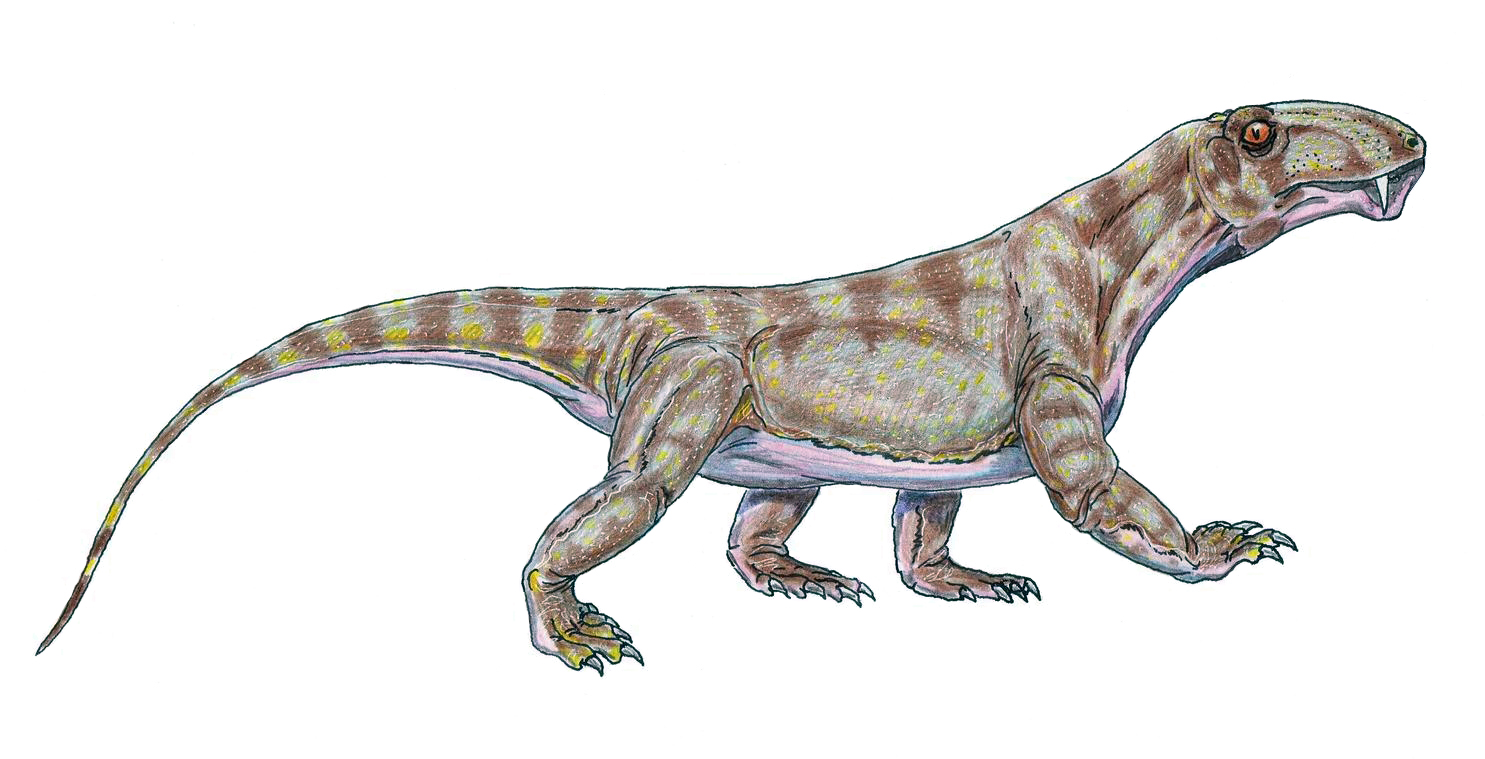
- Posture: Even for Hartman's 21st century Dimetrodon, there wasn't much difference between standing tall and sprawling flat. They didn't have to invest much energy in "standing up" before taking a step. Basal therapsids like Biarmosuchus (right) were different. With relatively short torsos, longer legs, and postures that brought the feet close to being underneath the body when they stood up, These were creatures that had to make a significant investment of energy in "standing up" before walking. One expects that, having made that investment, they would engage in longer bouts of walking around than their basal synapsid ancestors. That requires the energy we associate with tachymetabolism, at least during periods of activity.
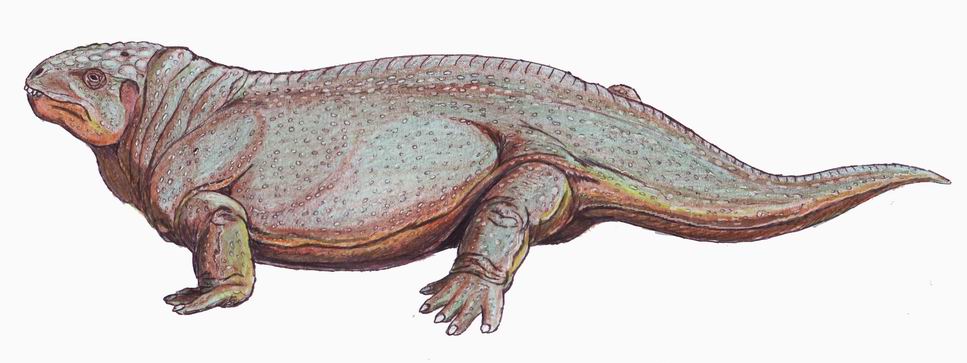
By analogy with living vertebrates, we would expect them to elevate their body temperatures during periods of activity, also. Alas, we lack good evidence for the condition of their glucose and oxygen plumbing, How elevated was their body temperature when they were active or at rest? Good question with only circumstantial evidence. Consider the "snuggling" Diictodon at right. Even when they share burrows (E.G. Youngina,) ectotherms tend not to snuggle to conserve body heat. What was Diictodon up to?
- Ventilation: When did the muscular diaphragm appear? One indication is the separation of the torso into distinct thoracic and abdominal regions. We begin to see the differentiation of dorsal vertebrae and ribs reflecting this at the base of Cynodontia.
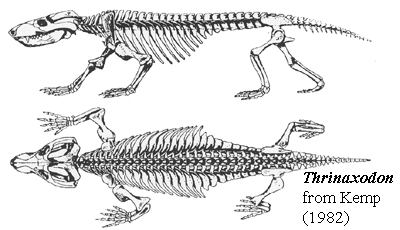
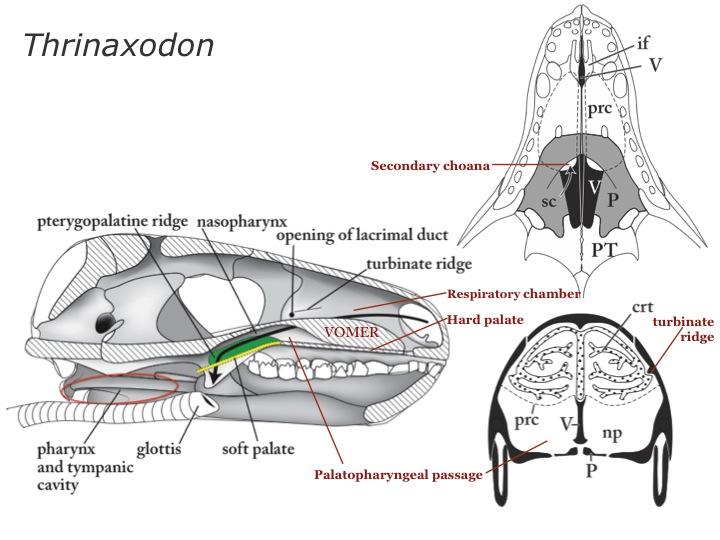
What about evaporative cooling mechanisms? Except for the mammaliaform Brasilitherium, only mammals have fully ossified nasal turbinates, however non-mammalian cynodonts and a few of their close relatives have ridges in their nasal cavities that seem to mark the attachment points for cartilagenous turbinates (see Thrinaxodon). All of this suggests an elevated ability ot ventilate the lungs among cynodonts.
- Fur: Alas, we depend on rare fossil occurrences. We know this much:
- Sail-back sphenacodontines would absolutely not have had it.
- Cynodonts similar to Morganucodon very close to the ancestry of mammals definitely had proper fur. (E.G.: Castorocauda fossil and reconstruction.)


One thing that does not occur until well after the common ancestry of proper Mammalia - a fully upright stance.
General Waypoints:
- Indications of homeothermy (indoor ectothermic):
- SA/Vol ratio
- Heat-exchange structures
- General indications of increased metabolic output:
- Anatomy suggesting prolonged bouts of locomotion
- Behavioral indictions of conservation of body heat
- Glucose-metabolism enhancements. Adaptations to rapid food processing:
- Grinding
- Shearing
- Ventilation enhancements - anything that moves more air through lungs
- Respiratory heat-exchange structures
- Integumentary structures
An Archosauriform scenario:
Assuming that archosauriforms followed an analogous evolutionary pathway, what stages of the acquisition of endothermy do the following taxa represent.
- Proterosuchidae:
- a tendency toward moderately large size
- blade-like serrated teeth
- sprawling stance
- Basal members of Eucrocopoda like Euparkeria:
- blade-like serrated teeth
- semi-erect stance, similar to basal therapsids.
- possible facultative bipedalism
- possible thoracic air cavities
- Loricata: Including Prestosuchus (right)
- Large size
- Hind limbs assume fully upright stance

Terrestrisuchus - a sphenosuchian-grade crocodylomorph - Sphenosuchian-grade crocodylomorphs: Including Terrestrisuchus (right)
- Small size
- Extremely slender long limbs.
- Modification of gastralia for use in belly-breathing
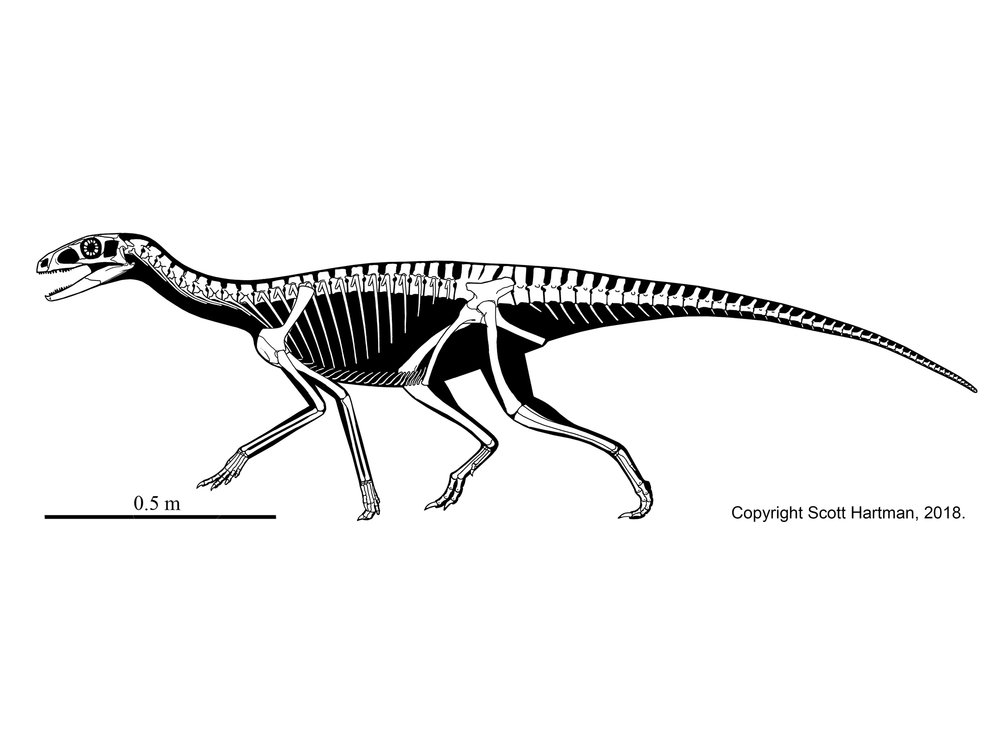
- Stem Dinosaurs: Including Silesaurus (right)
- Small size
- Extremely slender long limbs.
- Poposaurids: Including Effigia (right)
- medium size
- Obligate bipedalism.
- Basal Theropoda: Including Coelophysis (right)
- medium size
- Obligate bipedalism
- Extensive invasion of vertebrae by air-sac system
- Derived Theropoda: Including Velociraptor (right)
- small - medium size
- Obligate bipedalism
- Uncinate processes (odd doodads) on rib cage
- Anatomy suggests behaviors requiring agility and speed
- Close relatives with known integumentary structures
- Haversian bone
- Eusuchians: Including Bernessartia (right)
- medium - large size
- Fresh-water aquatic predators
- Alternating sprawling and semi-erect stance
- includes living Crocodylia
Caveat!
Because the thermoregulatory demands of terrestrial and marine life are so different, it is difficult to apply the same principles to them, however, based on isotope ratios, Bernard et al. 2010 determined that large ichthyosaurs, mosasaurs, and plesiosaurs regularly maintained high body temperatures, as well.








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}