
"God's noblest creation"
John Merck
And now, a problem....

Testudo, Malaclemys terrapin, the diamond-back terrapin

Stupendemys geographicus, the largest turtle ever
Pantestudines
(Permian - Quaternary) The total group containing Testudines and everything closer to them than to Lepidosauria or Archosauria. (Joyce et al., 2021.)No vertebrate group is more radically transformed than turtles - a situation that has greatly hampered attempts to ascertain their phylogenetic relationships. We have even complained of lacking the Archaeopteryx of turtles, despite, apparently, having at least one under our noses for a century. Here we start with what's definitely known and work outward.
Major groups
(Joyce et al., 2021.)- Pan-Testudines "The total clade of the crown clade Testudines" (In essence, Max ∇ (Testudines ~ Alligator, Lacerta))
- Testudinata "The clade for which a complete turtle shell, as inherited by Testudo graeca Linnaeus, 1758, is an apomorphy." (Effectively Min ∇ (Testudo, Proganochelys, Proterochersis))
- Testudines: Min ∇ (Chelonia, Trionyx, Chelonia, Testudo (cryptodires)). Includes:
- Cryptodira: The mid-line head-retracting turtles - Min ∇ (Trionyx, Kinosternon, Chelydra)
- Pleurodira: The side-necked turtles - Min ∇ (Chelus, Pelomedusa, Podocnemis)
Major trends:

Keratinous (above) and bony (below) elements of the shell from Pough et al., 1990.
- A dorsal carapace is integrated with a ventral plastron between the fore and hindlimbs. These sections form by distinct developmental processes and did not evolve simultaneously.
- Both carapace and plastron have:
- a bony endochondral skeleton
- a covering of broad keratinous scales.
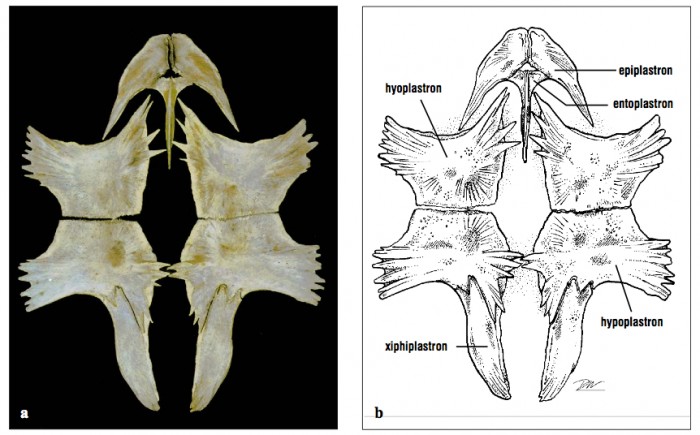
Elements in both layers are identifiable and have names. Bony and keratinous elements do not match up one for one.
Keratinous (left) and bony (right) elements of the shell in snapping turtle, Chelydra serpentina
- While the bones of the plastron are modified variants of those in other tetrapods:
- Epiplastron = clavicle
- Entoplastron = interclavicle
- Remainder = gastralia
The elements of the carapace appear to be composites of modified endochondral ribs and vertebrae, and dermal bone of some sort. The elements:
Ventral view of carapace of Apolone spinifera (on its back)- Neurals incorporate dorsal vertebrae
- Costals incorporate ribs
- Marginals, suprapygal, and pygal have no obvious homologs
- The nuchal's homologies are contentious (see below).
The turtle shell - Development:
How the components of the carapacial elements form and the precise identity of their homologs has been contentious. Major historic hypotheses include:
- The osteoderm hypothesis: maintains that the ribs and vertebrae fused developmentally with preexisting dermal scutes.
- The de novo hypothesis: holds that all of the shell elements are direct outgrowths of ribs and vertebrae.
in the costals:
- Ribs begin to form endochondrally, but are broadened by perichondral outgrowths.
- At the same time metaplastic (i.e. ectopic) bone forms in the adjacent dermis and fuses to the rib, yielding the platy costals.
Eventually, all suture to form a rigid shell. Thus, the development of the carapace is a little of both:
- The broadening of the ribs is absolutely a de novo process...
- But the metaplastic elements that fuse to them are, in essence, osteoderms.

Trachemys scripta, the red-eared slider - disassembled
- The ribs initially form in the somitic sclerotome but do not extend ventrally into the lateral plate mesoderm forming the body wall. Instead, they invade the dermis of the dorsal trunk.
- Rib formation is partially induced by the carapacial ridge that extends around the trunk. This feature functions like the apical ectodermal ridge of the limb buds, attracting the growing ribs and deflecting them from their normal developmental pathway. (Hirasawa et al., 2014)
- The elements of the plastron, in contrast seem to be greatly enlarged by a secondary proliferation of neural crest (Clark et al., 2001.) Moreover, the process that mediates plastron development seem, in addition, to suppress development of the cartilaginous sternum. (Rice et al., 2016)

Ventilating turtle lungs from Pough et al., 1990.
- The girdles are enclosed inside the shell. (Not strictly inside the rib cage when one disregards shell components that aren't strictly homologous with ribs.)
- There is no room for intercostal muscles normally used to expand and contract the rib cage in breathing.
- Breathing is facilitated by the action of muscles on the:
- posterior limiting menbrane posteriorly
- and on the pectoral girdle anteriorly.

Pectoral girdles of Captorhinus (left) and Macroclemys - a cryptodire (right)
- The turtle clavicle and interclavicle are incorporated into the plastron
- The cleithra are a puzzle:
- Bever et al., 2015 and Lyson et al. 2016 maintain that they are fused to form the nuchal plate of the carapace, whereas...
- Evidence for old-fashioned cleithra is reported for Kayentachelys (Joyce et al., 2006) and Eileanchelys (Anquetin et al., 2008)
- The pectoral girdle is triradiate with:
- a long, slender scapular shaft,
- a very long acromion process of the scapula,
- and a broad coracoid.
- the pelvis connects to the carapace through two pairs of sacral ribs, and abuts the plaston ventrally. (Among pleurodires, it fuses to the shell top and bottom.)

Chelydra serpentina, the snapping turtle - quadrate in red
- Marginal and palatal teeth lost
- Jaws lined by a rhamphotheca (keratinous beak) as in birds and dicynodonts
- The adductor muscles of the jaw pass over the otic capsule, which acts like a pulley.
- The basipterygoid articulation is fused. (Compare with Petrolacosaurus.)
- The skull is anapsid, yet there is often (but not always) significant emargination of the temporal skull roof by embayments from the occipital region and the lower cheek margin. (An extreme case.)

Diversity of Pantestudines
Testudines:
(Late Jurassic - Quaternary) The crown group of living turtles encompasses two distinct clades:- Cryptodira: the mid-line head-retracting turtles (familiar to North Americans)
- Pleurodira: the side-necked turtles (limited to the southern continents)
Synapomorphies of Testudines:
- Complete loss of palatal teeth
- Reduction of cervical ribs
- Reduction in tenth (posterior) dorsal rib
- Increase in number of pedal and manual phalanges


Chelodina longicollis the eastern long-necked turtle from Wikimedia.com (left) Terrapene carolina bauri the Florida box turtle of Gloria Lennon (right).
Pleurodira:
(side-neck - Late Jurassic - Quaternary) (above left)
Side-necked turtles retract their heads flexing their necks at the well developed articulations between cervical centra. Restricted to the southern (gondwanan) continents, they are the dominant turtles of Australia. Their synapomorphies include:

Comparison of stem testudine (left), cryptodiran (center) and pleurodiran trochlear pattern from Joyce, 2007
- Presence of a trochlear process of the pterygoid to guide the adductor muscles (right).
- Loss of the epipterygoid
- Presence of an anterior extension to the braincase wall
- Median contact of the posterior costals to form a suprapygal.
- Procoelous caudal vertebrae
- Pelvis solidly fused to shell
Pleurodiran superlatives:
- Stupendemys (Neogene of Venezuela) Fresh water aquatic. The largest turtle ever.
- Chelus fimbriata, the mata mata of South America. A cryptic turtle that has lost its beak and depends entirely on suction feeding.
- Chelodina longicollis, the Australian eastern long-necked turtle.
Cryptodira:
(Hidden neck - Late Jurassic - Quaternary) (above right)The monophyletic group containing all turtles that pull their heads straight into their shells. Synapomorphies include:
- Presence of an anterior extension to the braincase wall (convergent with Pleurodira!)
- Pterygoid - basisphenoid contact conceals basipterygoid articulation.
- Retraction of neck along sagittal plane facilitated by accessory articulations of the neural arches

- Paracryptodira: (Late Jurassic - Paleogene) Near the cryptodire/pleurodire split, including the common Baenidae (right). Abundant fresh water turtles of the Cretaceous and Paleogene.
- Chelonioidea: (Cretaceous - Quaternary) Sea turtles, the only turtles successfully to invade the open oceans, including the largest living turtle (the leatherback) and Cretaceous superlatives like Protostega.
- Trionychia: (Cretaceous - Quaternary) Including pig-nosed turtles and softshelled turtles - fresh water turtles with reduced carapaces and limbs modified for rapid swimming.
- Testudinoidea: (Paleogene - Quaternary) Including tortoises and pond turtles.

Closest Relatives of Testudines:
The fossil record is full of creatures that are clearly turtles for all intents and purposes but fall outside the Testudines crown. Starting at the crown we move downward.

Meiolania platyceps
- robust cranial horns and bosses
- long armored tails (independently derived)
Eileanchelys waldmani: (Middle Jurassic) Briefly noted - the first turtle with recognizable aquatic specializations.
Proganochelys: (Late Triassic) Turtles from farther down the tree are generally similar, with:

- Terrestrial adaptations (Illuminated by Joyce and Gauthier, 2004)
- Armored necks and tails

Proganochelys quenstedti
- Palatal teeth
- The absence of a trochlear system of the jaws
- A mobile basipterygoid articulation
- A clavicle (or is that "epiplastron?") with a dorsal process that reached the carapace.
- A relatively short acromion process.
Chinlechelys: (Late Traissic) Described by Joyce et al., 2008, the Late Triassic Chinlechelys of Arizona resembles Proganochelys except in that perichondral bone of its ribs and vertebrae are so loosely coossified with their dermal component, that their separate identity is clear. When first described, this seemed to resolve a long-standing enigma. In this early testudinatan, the skeletal elements of the shell seem to form by the coossification of the ribs with preexisting dermal plates, but it was not to be.
Testudinata: (Late Triassic - Quaternary) The last common ancestor of all creatures with complete turtle shells, and all of its descendants. Synapomorphies include:
- Ten dorsal vertebrate present
- Costal elements sutured together
- Nuchal, pygal, and costal elements present
Stem Testudinata: Now it gets interesting.

Odontochelys semitestacea
Odontochelys semitestacea:
Described by Li et al., 2008, the Late Triassic Odontochelys of China was at the time the most anatomically primitive turtle known. Indeed, it definitely lacks one synapomorphy - the toothless mouth. Odontochelys has typical simple teeth. Stranger yet, although it has a plastron, it lacks a proper carapace.Distinctive features:
- Its depositional environment and limb shape indicates that it was aquatic.
- It's ribs are expanded into paddle-shapes, but
- there are no endochondral dermal scutes associated.
- Some living highly aquatic turtles have reduced their carapaces significantly.
- However the plastron forms from a secondary proliferation of neural crest, a distinct developmental pathway from the carapace.
Synapomorphies with Testudinata:

- Paired gastralia transformed into plastron (with addition of metaplastic bone)
- Cartilaginous sternum is lost (plastron osteogenic process underway)
- Addition of two cervical vertebrae
- incipient development of dorsal vertebrae into neural plates
- Ribs are broadened and T-shaped in cross-section. Adjacent ribs abut one another but are not co-ossified.
- Tips of ribs appear to invade the dermis developmentally - carapacial ridge present.
Odontochelys was found in marine deposits of an embayment that was surrounded on three sides by land. Was it aquatic? That was the initial interpretation of Li et al., 2008 however it:

- is discordant with other animals in this part of the tree
- is not strongly supported anatomically. Morphometric comparative analyses by Joyce, 2015 and Dudgeon et al., 2021 maintain that the morphology of forelimbs in Odontochelys and other stem testudinatans supports terrestrially with, at most, occasional trips into the water.

Eorhynchochelys sinensis:
Li et al., 2018 Provide a glimpse of a slightly less derived version from the Early Late Triassic of China. Eorhynchochelys has the broadened ribs and flattened puboischiatic plate of turtles, but lacks any obvious plastron, offering a glimpse of the pantestudine condition just prior to that transformation. Caution, though - this region of the body is poorly exposed in the fossil.
But wait! Eorhynchochelys is derived in one respect: Although it retains maxillary teeth, the premaxillae and anterior dentaries seem to support a rhamphotheca!

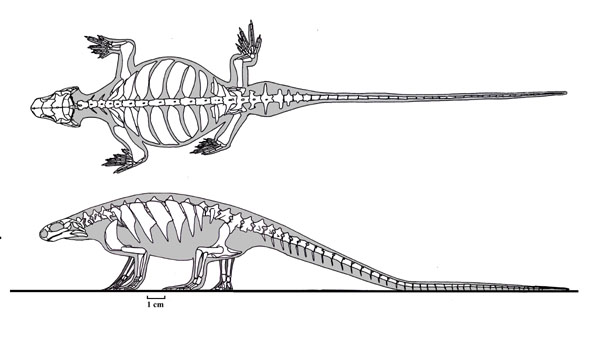
Eunotosaurus africanus:
For a century, we have lamented not having the Archaeopteryx of turtles despite, evidently actually having it. Eunotosaurus africanus: (Middle Permian) Roughly a foot long, with a small head and roly-poly torso supported by expanded dorsal ribs. Known to science since 1892, this animal has alternately been considered close to and far from turtle ancestry.Eunotosaurus shares with Odontochelys and Testudinata these synapomorphies:
- Reduced number of dorsal vertebrae
- Dorsal vertebrae elongate compared to cervicals and sacrals
- Dorsal ribs perichondrally expanded
- Judging from histology, intercostal muscles did not span the gaps between dorsal ribs.
- Trunk is wide
- Paired gastralia (i.e. missing the typical mid-line gastrale, but otherwise normal-looking)
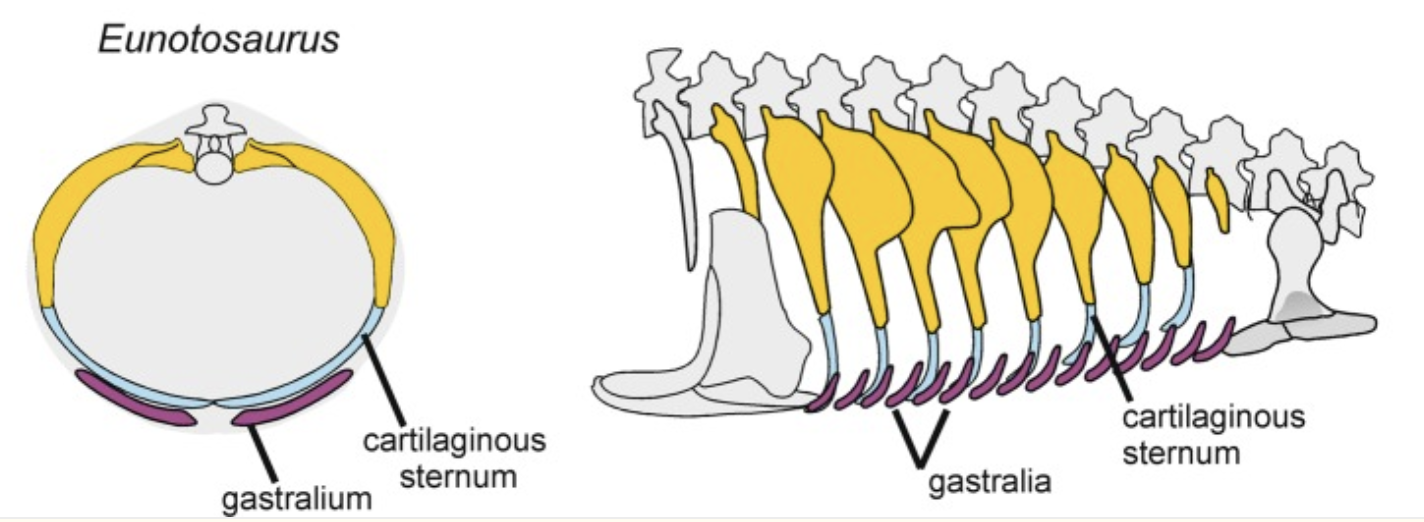
Eunotosaurus africanus thorax schematic from Schoch et al., 2019
(BTW, it's not "gastralium," it's "gastrale.")
But let's be clear about how Eunotosaurus is plesiomorphic:
- Ribs, while overlapping slightly, have some mobility relative to one another.
- They develop as normal ribs do in the lateral plate mesoderm. There is no evidence of a carapacial ridge.
- No evidence of metaplastic ossifications of the dermis. It's just ribs.
- Finally, all evidence suggests the continued presence of a cartilaginous sternum, as in other amniotes.
So, if Eunotosaurus' broad ribs didn't confer protection, what were they good for, especially considering that they reduce stride length by making the trunk inflexible? Lyson et al., 2016 argue that they are an adaptation to fossoriality, providing Eunotosaurus, with a solid base for the origin of powerful limb muscles. This is in line with other limb-digging fossorial vertebrates like Thrinaxodon.
Regardless of how phylogenetic analyses are structured, Eunotosaurus groups with Odontochelys and Testudinata.


Caveat:
But note, in some ways Eunotosaurus really isn't what we expect of saurian or nearly saurian turtle precursor:- Its rod-shaped squamosal is reminiscent of that of a procolophonoid ankyramorph and is dissimilar to the four-pronged squamosals of Parapleurota
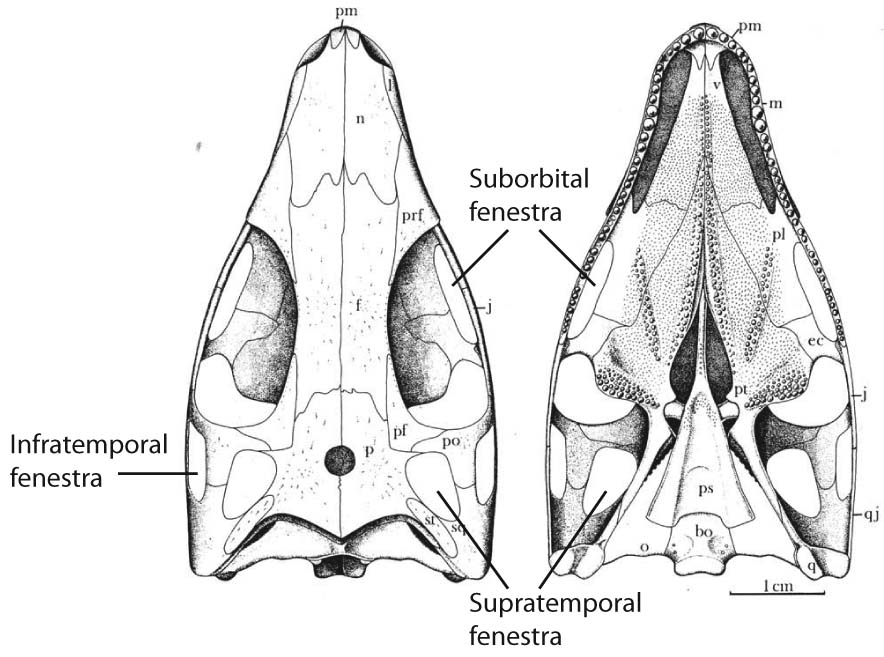
- It lacks a suborbital fenestra in the palate
- Its ribs imbricate with one another (overlap) in a manner unlike other stem testudinatans
Ontogeny and Phylogeny
One story that emerges from this phylogeny involves the position of the pectoral girdle with respect to the ribs. Ontogenetically, the scapula first forms in the normal position, but is able to slip beneath the carapace because the anterior ribs become angled posteriorly. Phylogenetically we see the same sequence:- In Eunotosaurus the scapular blade is oriented dorsally anterior to the rib cage.
- In Odontochelys the dorsal process of the scapula is short and well anterior to the ribs because the distal ends of the ribs are squeezed together anteroposteriorly
- In Proganochelys the ribs (as costals) have resumed their normal orientation but the scapula is now ventral to them.

The Position of Pantestudines - May, 2015
Follow the bouncing turtles: When we try to say anything about the position of turtles in the tree of amniote evolution, things quickly get weird. Of all recent phylogenetic enigmas, turtles have been the most intractable.Chronology of phylogenetic hypotheses:
- Gaffney and Meylan, 1988, deemed them to be related to captorhinids.
- Reisz and Laurin, 1991 indicated a possible close relationship between turtles and procolophonoids.
- Lee, 1993 proposed Pareiasaurs as a sister taxon. (In fact, according to later work by Lee, Pareiasaurs may be paraphylteic with respect to turtles, i.e. turtles may be a kind of pareiasaur.) Lee's argument assumed that the dermal component of the turtle shell derived from preexisting osteoderms.
- Rieppel and deBraga, 1996 performed a morphological cladistic analysis that placed turtles inside Lepidosauria, as members of Euryapsida, alongside sauropterygians. An interesting result for what it says about euryapsids! Traditionally turtle armor is seen as fundamentally different from that of armored placodonts but this result prompted open reconsideration.
- Lyson et al., 2010 re-ran the Rieppel and deBraga 1996 matrix adding Eunotosaurus and Proganochelys. The effect was that Eunotosaurus grouped as the sister taxon of Testudinata, inside Parareptilia.
Data type and results
Molecular analyses: For twenty years, relationships of Testudines to other extant groups have been addressed by molecular systematics. At various points they have been recovered as sister taxon to:- Sauria
- Lepidosauria
- Archosauria
- Crocodylia
- Aves
2015 - Summer of Revelation:
When we taught GEOL431 in 2015, the accumulating morphological and developmental data seem to support the ankyramorph hypothesis of turtle origins. Then came the great enlightenment:
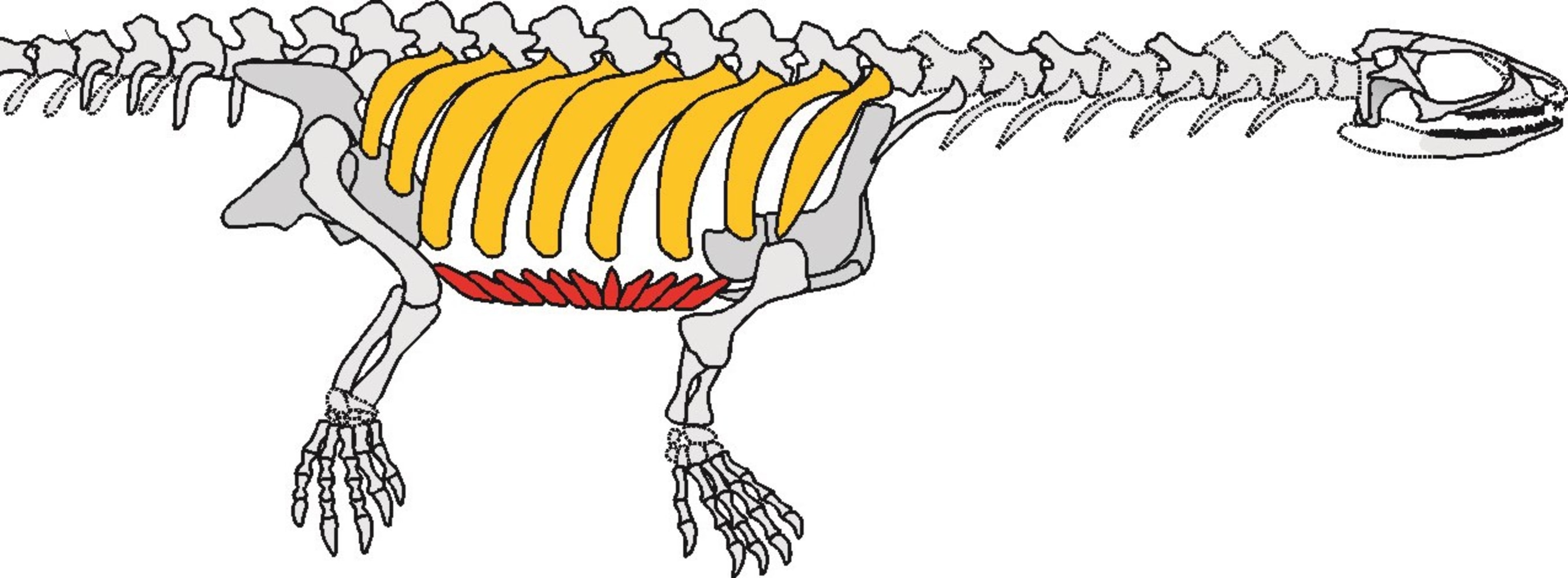
Pappochelys rosinae (Middle Triassic of Germany) was described by Schoch and Sues, 2015. This small (~30 cm overall length) animal is intermediate between Eunotosaurus and Odontochelys both in time and morphology:
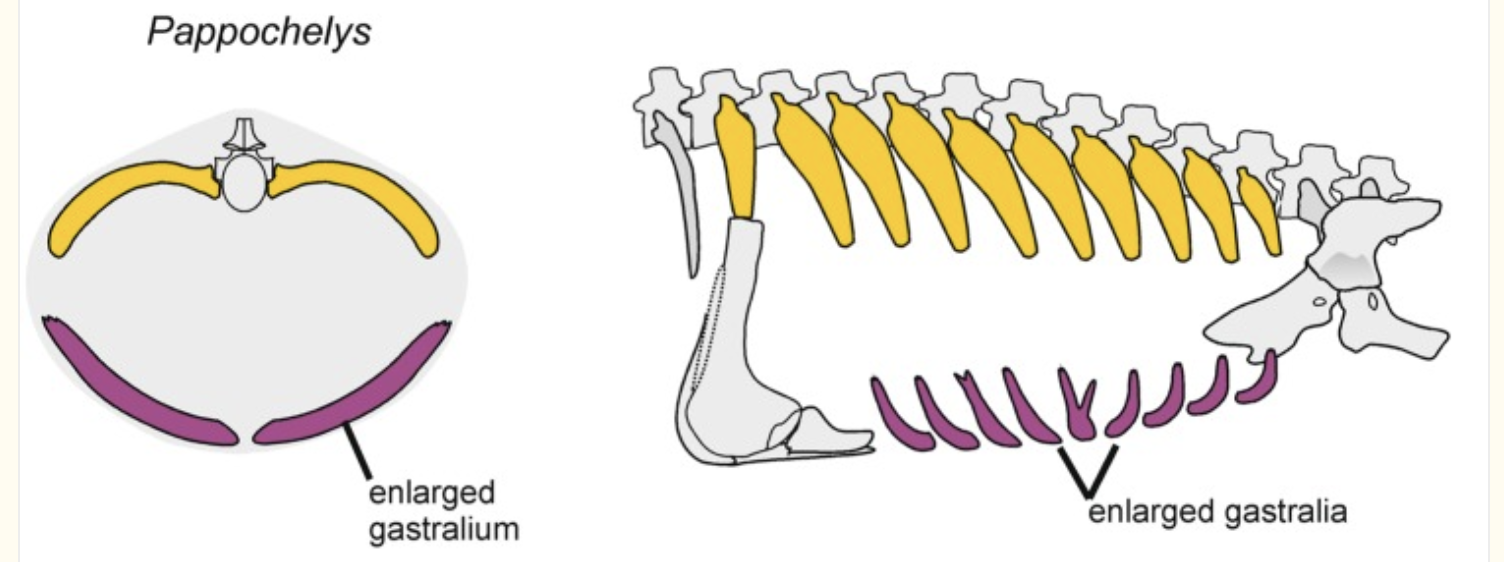
Pappochelys trunk schematic from from Schoch et al., 2019
(BTW, it's not "gastralium," it's "gastrale.")
- Tips of ribs appear to invade dermis developmentally - carapacial ridge!
- Its ribs are expanded but non-overlapping. No evidence of metaplastic bone.
- Its gastralia are very robust - sometimes branching distally - but not developed into a plastron.
- The cartilaginous sternum appears to be suppressed.
But its remarkable feature is its diapsid skull, that with its ventrally open infratemporal fenestra resembles that of a derived stem saurian. It appears that turtles are, in fact, some kind of parapleurotan (crownward diapsid.)
But what about Eunotosaurus? It has an anapsid skull, right?
Not actually. The skull roof of Eunotosaurus is covered with dermal bone, however it has an odd feature: the supratemporal projects far anteriorly. By digitally removing it form high-resolution CT scans, Bever et al., 2015 revealed that it covered a proper supratemporal fenestra. Moreover, they showed ontogenetic variation in the development of the anterior extension of the supratemporal bone. Juveniles, it developed, had proper supratemporal fenestrae.

{kind=link}
{kind=link}
.png){kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Why would Eunotosaurus do this?
- The strengthening of the skull is common in fossorial animals. Seeing this in Eunotosaurus is consilient with Lyson et al.'s, 2016 argument for fossoriality.
And what does this tell us about the importance of fenestration patterns as "key characters" in taxonomy? :-(
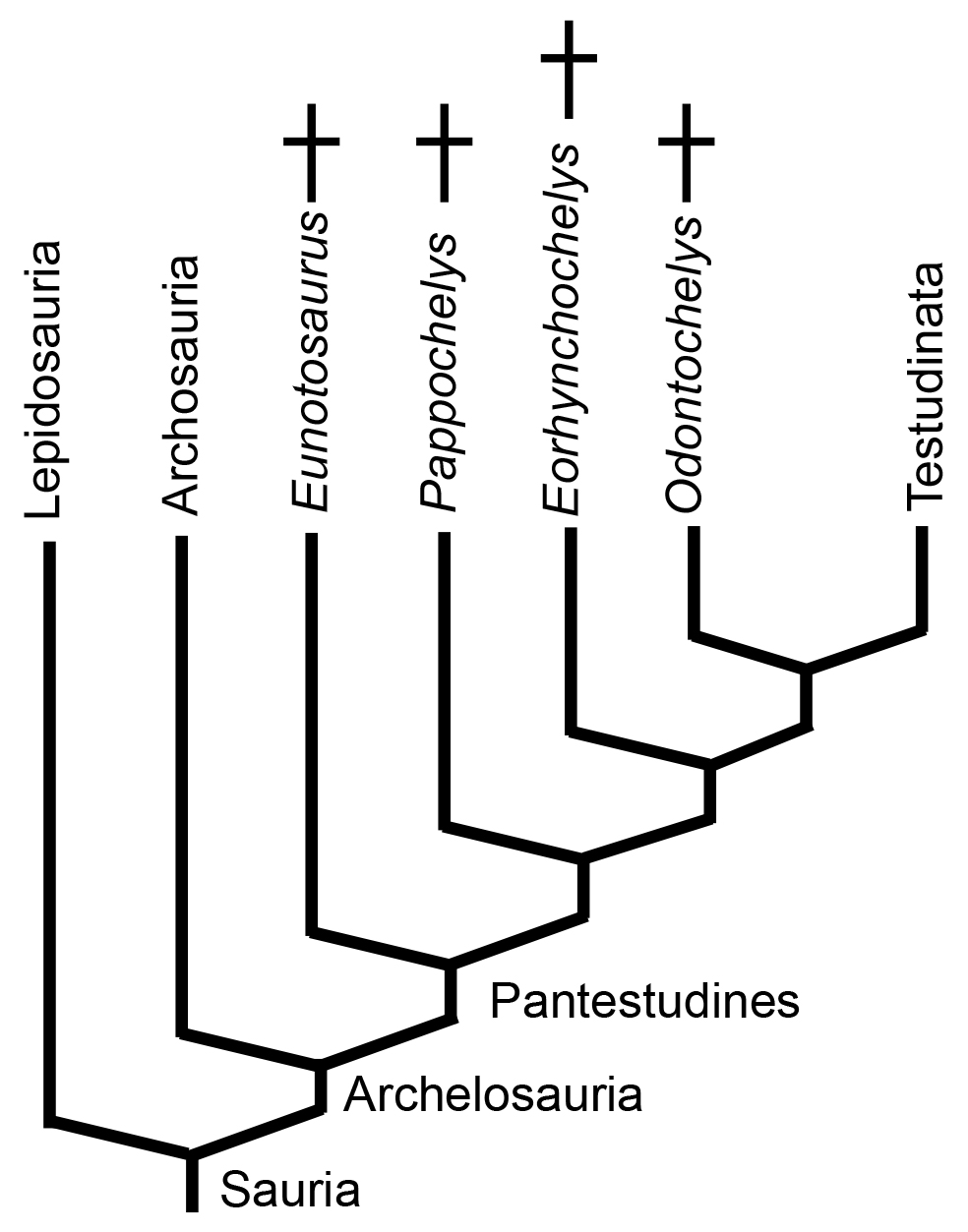
Pantestudines as of Summer, 2025. The end?
So what are they?
As of 2025, the weight of evidence and opinion supports Pantestudines as a basal branch of Archosauromoprha. Together with Archosauria, they form the clade Archelosauria (Crawford et al., 2012). Your instructors can live with this, especially as Simões et al. 2022 have identified potential morphological synapomorphies with Archosaurs:- Sagittal crest of supra occipital
- Absence of ectepicondylar foramen of humerus.
An yet....
Pappochelys is informally reported (Sues, 2015 pers. comm.) to have:
- a facet for the cleithrum on its scapula. Moreover, evidence for cleithra is reported for Kayentachelys (Joyce et al., 2006) and Eileanchelys (Anquetin et al., 2008) as well.
- no descending flanges of the parietal
That indicates that although they may be diapsids, they are outside of Sauria.
We hope for salvation through future revelations.
Amen.
- Anquetin, J., Barrett, P.M., Jones, M.E.H., Moore-Fay, S., and Evans, S. 2008. A new stem turtle from the Middle Jurassic of Scotland: new insights into the evolution and palaeoecology of basal turtles. Proceedings of the Royal Society B, 276:879-886. https://doi.org/10.1098/rspb.2008.1429
- G. S. Bever, Tyler R. Lyson, Daniel J. Field, and Bhart-Anjan S. Bhullar, 2015. Evolutionary origin of the turtle skull. Nature 525: 239-242.
- Juan C. Cisneros, Ross Damiani, Cesar Schultz, Atila da Rosa, Cibele Schwanke, Leopoldo W. Neto and Pedro L. P. Aurelio, 2004. A procolophonoid reptile with temporal fenestration from the Middle Triassic of Brazil. Proceedings of the Royal Society B, July 2004.
- K. Clark, G. Bender, B.P. Murray, K. Panfilio, R. Davis, K. Mumen, R.S. Tuan, S. F. Gilbert, 2001. Evidence of the neural crest origin of turtle plastron bones. Genesis 31(3): 111-117.
- Nicholas G. Crawford, James F. Parham, Anna B. Sellas, Brant C. Faircloth, Travis C. Glenn, Theodore J. Papenfuss, James B. Henderson, Madison H. Hansen, W. Brian Simison, 2015. A phylogenomic analysis of turtles, Molecular Phylogenetics and Evolution 83, 2015, 250-257,ISSN 1055-7903, https://doi.org/10.1016/j.ympev.2014.10.021.
- Thomas W. Dudgeon, Marissa C. H. Livius, Noel Alfonso, Stephanie Tessier, Jordan C. Mallon, 2021. A new model of forelimb ecomorphology for predicting the ancient habitats of fossil turtles. Ecology and Evolution 11(23) 17071-17079.
- Eugene Gaffney, 1990. The comparative osteology of the Triassic turtle Proganochelys. Bulletin of the American Museum of Natural History. 194. 261 pp.
- Eugene Gaffney and Peter Meylan, 1988. A phylogeny of turtles. pp. 157-219 in M.J. Benton (ed.) The Phylogeny and Classification of the Tetrapods, Vol.1, Clarendon Press, Oxford, 1988.
- Tatsuya Hirasawa, H Nagashima, Shigeru Kuratani, 2013. The endoskeletal origin of the turtle carapace. Nature Communications. 4: 2107.
- Tatsuya Hirasawa, Juan Pascual-Anaya, Naoki Kamezaki, Mari Taniguchi, Kanako Mine, and Shigeru Kuratani, 2014. The evolutionary origin of the turtle shell and its dependence on the axial arrest of the embryonic rib cage. Journal of Experimental Zoology (Mol. Dev. Evol.) 324B: 194-207.
- Walter Joyce and Jacques Gauthier, 2004. Palaeoecology of triassic stem turtles sheds new light on turtle origins. Proceedings in Biological Science 7:271(1534):1-5.
- Joyce, W.G., Jenkins, F.A., and Rowe, T.R. 2006. The presence of cleithra in the basal turtle Kayentachelys aprix. Fossil Turtle Research, 1:93-103.
- Walter Joyce, 2007. Phylogenetic relationships of Mesozoic turtles. Bulletin of the Paebody Museum of Natural History 48(1): 3-102.
- Walter Joyce, Spencer Lucas, Torsten Scheyer, Andrew Heckert, and Adrian Hunt, 2009. A thin-shelled reptile from the Late Triassic of North America and the origin of the turtle shell. Proceedings of the Royal Society B.276: 507-513.
- Walter Joyce, 2015. The origin of turtles: A paleontological perspective. Journal of Experimental Zoology (Mol. Dev. Evol.) 324B: 181-193.
- Walter G. Joyce, Jeremy Anquetin, Edwin-Alberto Cadena, Julien Claude, Igor G. Danilov, Serjoscha W. Evers, Gabriel S. Ferreira, Andrew D. Gentry, Georgios L. Georgalis, Tyler R. Lyson, Adán Pérez-García, Márton Rabi, Juliana Sterli, Natasha S. Vitek and James F. Parham, 2021. A nomenclature for fossil and living turtles using phylogenetically defined clade names. Swiss Journal of Palaeontology, 140: 5.
- Michael Lee, 1993. The origin of the turtle body plan: bridging a famous morphological gap. Science 261: 1716-1720.
- Chun Li, Nicholas C. Fraser, Olivier Rieppel, and Xiao-Chun Wu, 2018. A Triassic stem turtle with an edentulous beak. Nature 560, 476–479.
- Chun Li, Xiao-Chun Wu, Olivier Rieppel, Li-Ting Wang, and Li-Jun Zhao, 2009. an ancestral turtle from the Late Triassic of southwestern China. Nature 456/27.
- Asher J. Lichtig and Spencer G. Lucas, 2021. Chinlechelys from the Upper Triassic of New Mexico, USA, and the origin of turtles. Palaeontologica Electronica: 24.1.a13.
- Tyler Lyson, Gabe Bever, Bhart-Anjan Bhuller, Walter Joyce, and Jacques Gauthier, 2010. Transitional fossils and the origin of turtles. Biology Letters 6, 830-833.
- Tyler Lyson, Gabe Bever, Torsten Scheyer, Allison Hsiang, and Jacques Gauthier, 2013. Evolutionary Origin of the Turtle Shell. Current Biology 23, 1113-1119.
- Tyler Lyson, Bruce S. Rubidge, Torsten M. Scheyer, Kevin de Queiroz, Emma R. Schachner, Roger M.H. Smith, Jennifer Botha-Brink, G.S. Bever, 2016. Fossorial Origin of the Turtle Shell. Current Biology 26, 1887-1894.
- Sean Modesto, 2000. Eunotosaurus africanus and the Gondwanan ancestry of anapsid reptiles. Palaeontologia Africana 36: 15–20.
- Robert Reisz and Michel Laurin, 1991. Owenetta and the origin of turtles. Nature 349:324-326.
- Ritva Rice, Aki Kallonen, Judith Cebra-Thomas, and Scott F. Gilbert, 2016. Development of the turtle plastron, the order-defining skeletal structure. Proceedings of the National Academy of Sciences April 25, 2016 113 (19) 5317-5322
- Olivier Rieppel and Michael DeBraga, 1996. Turtles as diapsid reptiles.Nature 384: 453-455.
- Rainer Schoch and Hans-Dieter Sues, 2015. A Middle Triassic stem-turtle and the evolution of the turtle body plan. Nature 523: 584-587.
- Rainer R. Schoch, Hans-Dieter Sues, 2019. The origin of the turtle body plan: evidence from fossils and embryos. Palaeontology 63(3) 375-393.
- Rainer R Schoch, Nicole Klein, Torsten M Scheyer, Hans-Dieter Sues, 2019. Microanatomy of the stem-turtle Pappochelys rosinae indicates a predominantly fossorial mode of life and clarifies early steps in the evolution of the shell. Sci Rep. 2019 Jul 18;9(1):10430. doi: 10.1038/s41598-019-46762-z. PMID: 31320733; PMCID: PMC6639533..
- Simões TR, Kammerer CF, Caldwell MW, Pierce SE. Successive climate crises in the deep past drove the early evolution and radiation of reptiles. Sci Adv. 2022 Aug 19;8(33):eabq1898. doi: 10.1126/sciadv.abq1898. Epub 2022 Aug 19. PMID: 35984885; PMCID: PMC9390993.
- Ingmar Werneburg, Wolfgang Maier, Walter Joyce, 2013. Embryonic remnants of intercentra and cervical ribs in turtles. Biology Open 000: 1-5.
- Ingmar Werneburg and Marcelo Sanchez-Villagra, 2009. Timing of organogenesis support basal position of turtles in the amniote tree of life. Evolutionary Biology 9:82.
- G. S. Bever, Tyler R. Lyson, Daniel J. Field, and Bhart-Anjan S. Bhullar, 2015. Evolutionary origin of the turtle skull. Nature 525: 239-242.