
Fossil Jawless Vertebrates
Stem Vertebrates:
Cambrian lagersäthen, mostly Chengjiang and the Burgess Shale provide early glimpses of the first craniate chordates - stem vertebrates.Myllokunmingia and Haikouichthys of Chengjiang give us the best picture picture of what the ancestral vertebrate might have looked like with Haikouichthys ercaicunensis (right - possibly junior synonym of Myllokunmingia fengjiaoa) (Shu et al., 1999) which seem to preserve:

- W-shaped myomeres
- placode-derived special sense organs
- a branchial skeleton resembling that of lampreys
- Dorsal fin, with anteriorly sloping linear texture.
- paired cartilaginous elements near the anterior notochord
They are not alone.
A short list of Early - Middle Cambrian soft-tissue stem vertebrates has arisen over the last decade, including:

- Nuucichthys of the Marjum Fm. Of Utah. (Lerosey-Aubril and Ortega-Hernandez, 2024.)
- Metaspriggina: of the Burgess Shale. (Conway-Morris and Caron, 2014)
- internal to the gills
- Each branchial arch has an upper and lower element

Yunnanozoon
No consensus exists on their exact branching sequences, however:
- If Yunnanozoon is a stem vertebrate, it branched off before the evolution of the head and special sense organs.
- Only Myllokunmingia and Haikouichthys possess anything resembling a dorsal fin supported by fin-rays. The median fins (dorsal and anal), according to Bailon-Zambrano et al., 2024, arise from expansion of the sclerotome. (In contrast, the median fins of Branchiostoma follow different developmental pathways.) Did Nuucichthys and Metaspriggina lack median fins because they had not yet evolved this sclerotome expansion before branching off?
Hints of unseen diversity - conodonts

Conodont elements
- Highly diverse and rapidly evolving, thus excellent index fossils.
- Originally proposed to be the teeth of some unknown fish, but paleontologists soon determined they were were clueless about:
- What kind of animal they were from
- What part of the animal they represented.
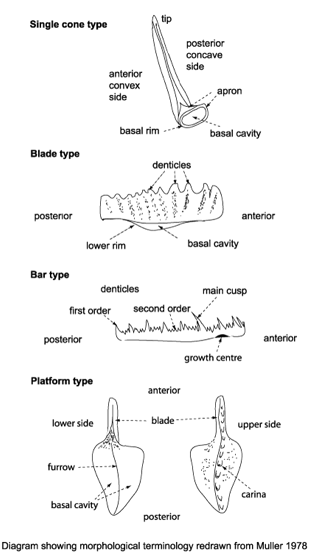
Conodont types
At this point, speculation raged about:
- Which major taxon the elements belonged to
- How were they used? Originally proposed to be dental elements of fish-like chordates by Pander (1856), but soon, more erudite-sounding hypotheses held sway.
- To some they seemed likely to be used in prey capture, as in similar structures in chaetognaths (arrow-worms)
- To others, they seemed like mineralizations of the suspension feeding apparatus of cephalochordates.
- Or perhaps they were for internal support and not for feeding at all.

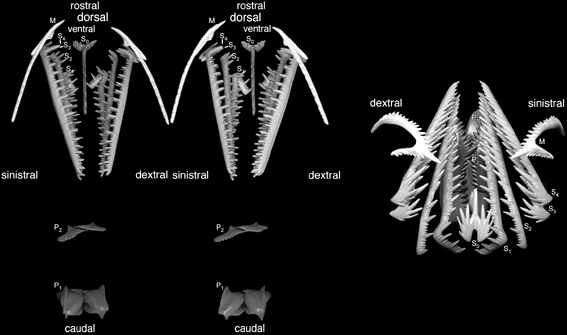
Conodont apparatus
Moreover, different elements began to be distinguished by the manner in which they were formed developmentally.
Since these seemed to appear in a stratigraphic (chronological) sequence, speculation was that these growth forms represented evolutionary stages. Not quite. In 1988, single cone elements were eliminated from the roster of euconodonts by the discovery that they were the fossils of early chaetognaths. In living chaetognaths, similar elements are made of calcium carbonate and are used in prey capture.
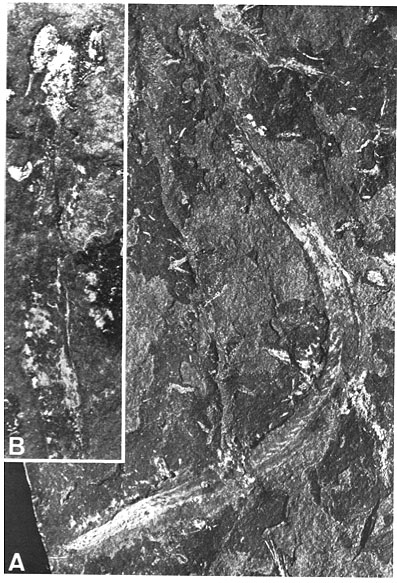
Clydagnathus
- Chordate-like V-shaped segmented muscle blocks
- Midline fins supported by fin rays
- The conodont apparatus in an anterior position, suitable for use in feeding.
- Notochord
- A head a brain and two capsules for special senses, thought to be very large eyes and smaller otic capsules.
More illumination came from the very well-preserved Promissum pulchrum, a 40 cm. euconodont (Gabbott et al. 1995). It shows V-shaped (not W-shaped) myomeres and evidence for extrinsic eye muscles.

Euconodonts
But where does Euconodonta go on the chordate cladogram? The presence of phosphatic hard parts arguably places it, closer to Gnathostomata than to hagfish or lampreys, but there are concerns:
- The conodont elements, as elements, are not homologous to any other phosphatic vertebrate feature.
- Jawless vertebrates might have secondarily lost hard tissues as a reversal.
- Some morphological interpretations of euconodonts, especially the huge eyes, seem to cry out for revision.
- The assumption of euconodont monophyly hasn't really been rigorously tested.
The vertebrate evolutionary pattern:

We have such a copious record of heavily armored Early Paleozoic forms that it is tempting to forget that the group's most basal members (like Myllokunmingia) essentially lacked hard tissues. Indeed, the early evolution of Vertebrata is marked by the diversification of bony tissues and their proliferation through the body. This pattern is illuminated by Sansom et al., 2010 and Miyashita et al., 2019. So, we start with a review of bony evolution as revealed in these studies.
Note: The taxa depicted fall into three broad categories:
- Basal boneless craniates (Myllokunmingia, Metaspriggina, etc.)
- Total group Cyclostomi (Including Euconodonta!)
- Total group Gnathostomata

Bones
Hagfish and lampreys, as the only living jawless vertebrates, provide an interesting glimpse of early vertebrate evolution, however they lack the proper hard tissues by which we know the vast diversity of early vertebrates, including bone.
Fossil vertebrates are mostly known from hard tissues - bone and teeth. Bone is composed of:
- A mineral component - made of calcium phosphate (i.e. the mineral hydroxyapatite).
- A protein component - made mostly of the fibrous protein collagen.
- Osteoblasts: Active cells that secrete bone's extracellular matrix
- Osteocytes: When osteoblasts become trapped inside the bony matrix, they become inactive osteocytes and function in maintenance. In both life stages, these cells extend long processes (extensions) through canals in the bone. Osteoblast/osteocytes may be derived from nerve cells, whose axons and dendrites are similar.
(Note: The corresponding cells responsible for bony tissue in teeth are called:
- odontoblasts/odontocytes when they secrete dentin
- ameloblasts when they secrete enamel)
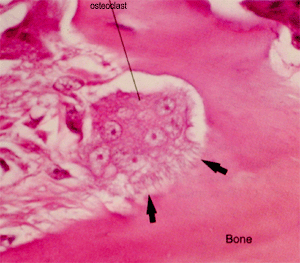
- Osteoclasts: play a major role in bone remodeling by removing bone. They resemble and might be derived from macrophages.
Bone's potential functions include:
- Skeletal support for muscular system
- Armor
- Reservoir of phosphate ion - a scarce and limiting biological material.
Bony tissue can be:

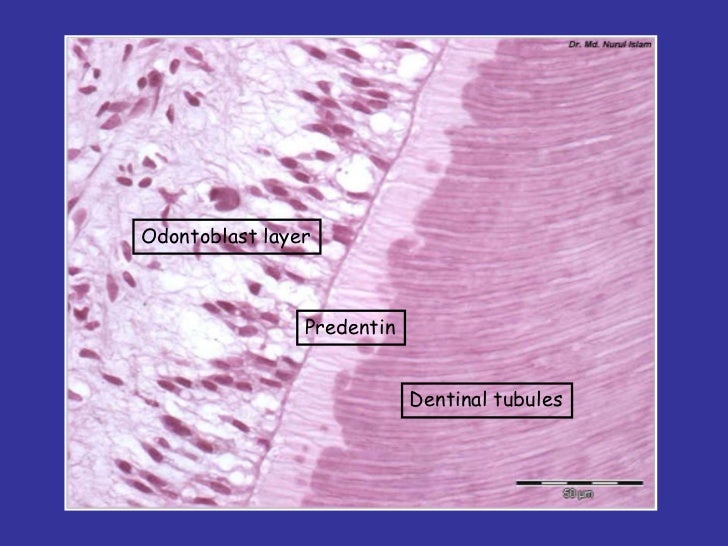
- Acellular if the bone cells migrate away as the bone is laid down and not enclosed by it. Particularly dense acellular bone (~90% mineral) is dentin (aka "dentine"), characteristic of teeth and placoid scales. Dentin is characterized by parallel dentin tubules housing processes of the retreating odontoblasts.
- Cellular if the bone cells are enclosed inside the bone (right).
What is its history?
The earliest known phosphatic hard tissues were acellular, and were tooth-like in being made of:
- Dentin: Dense acellular bone
- Enamel: Bony tissue made entirely of interlocking crystals of hydroxyapatite (i.e. lacking collagen). Note: Specialists distinguish enamel, secreted entirely by ameloblasts, from enameloid in which both ameloblasts and odontoblasts (or analogous cells) participate. For details, see Houée et al., 2024.
Bone in any form only occurs among members of Vertebrata. What does the study of fossil organisms tell us about the distribution of bony tissue?
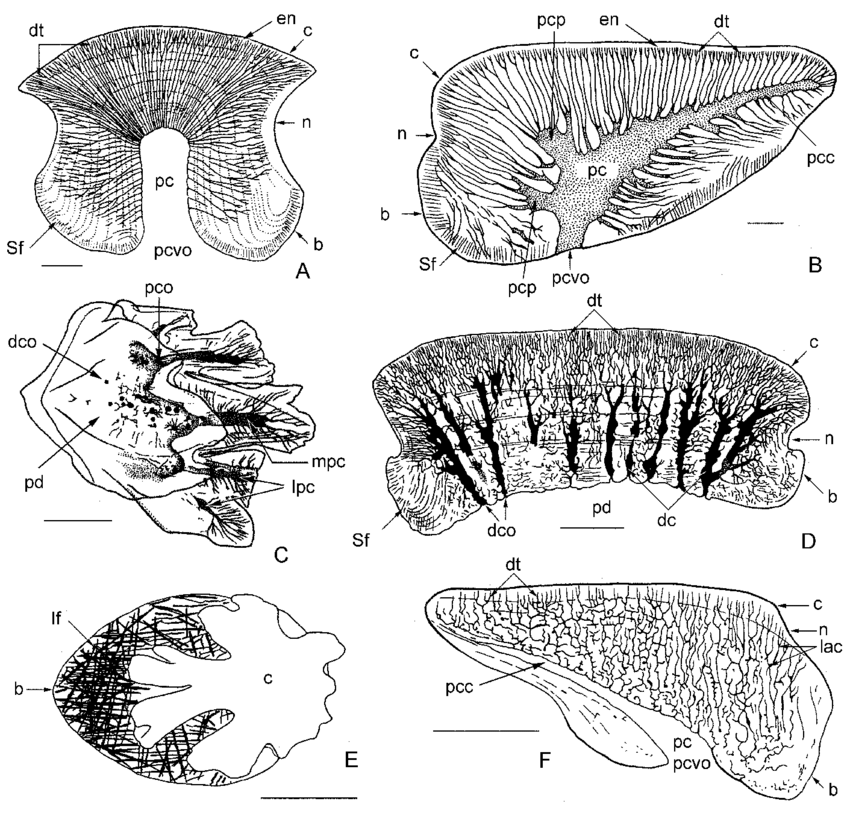
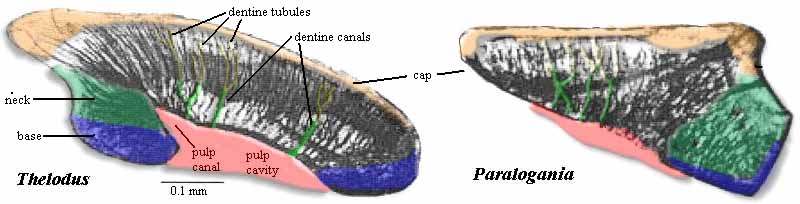
Thelodont scales - basic vertebrate denticles with pulp, dentin, and enameloid.
A rogue's gallery of early Paleozoic vertebrates:
As we do this survey, be aware of these issues:
- The proliferation of bone in the body
- The rise of the diversity of types of bony tissue
- Changes in the feeding apparatus
- Changes in the organization of the nasal capsules, hypophysis, and nasohypophyseal duct.
- Appearance of fin-like structures
As shown at right, a fundamental unit of vertebrate hard tissue took the form of denticles with pulp-cavities, dentin, and enameloid.

Anatolepis armor
The most basal vertebrates, however, lacked any hard tissues (except for conodont elements.) A survey of early vertebrate evolution should focus on their acquisition:
Total Group Cyclostomi
- Hypocercal tail
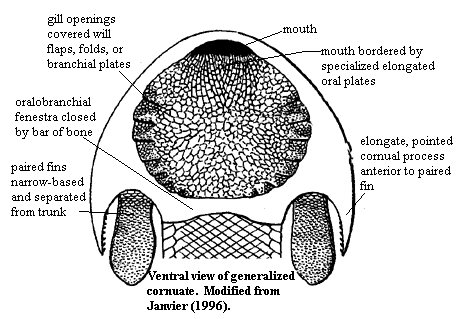
- Anywhere from six to thirty branchial (gill) openings (compared to seven for lamprey). These usually formed a posteroventrally slanting row. In exceptionally preserved specimens of Jamoytius and Euphanerops, gills are organized in pouches with external branchial cartilage, like in lamprey.
- Retain a single median nasohypophyseal opening and nasal capsule (again reminiscent of lampreys).
- Single anal fin
- Lateral fin folds: hagfish and lampreys have no suggestion of paired appendages. In Euphanerops, Jamoytius, Lasanius, and Anaspida, however a pair of long lateral pleats extend down the ventral margins of the animal's trunk. They contain neither cartilage nor muscle, but suggest a starting point for the evolution of paired appendages - a house waiting to be inhabited by appendicular muscles and skeleton. Note: Although they lack paired appendages, lampreys do have distinct lateral plate somatic mesoderm (Onai, 2023.)
- In anaspids, there are, additionally, paired arrays of triangular spines (1 - 6) in a vaguely "pectoral" position. (See Reeves et al., 2023 for details of Lasanius.)
- Hard tissue takes the form of acellular strap and plate-like scales. In Jaymoutius and Euphanerops, these only occur on the trunk, but in Anapsida they extend onto the head.
- Dermal skeleton.
- Anal fin
- Paired fin folds

Pharygolepis, an anapsid
Anaspida (Silurian), Euphanerops (Devonian), and Jamoytius (Silurian)
These form a clade in analysis of Miyashita et al., 2019. Illuminated by well-preserved Jamoytius, Lasanius, and Euphanerops. These were cylindrical, vaguely lamprey-like creatures with varying amounts of acellular dermal bone plates and scales, but without specialize mouthparts. They lack obvious adaptations to suspension feeding or to taking large prey. Deposit feeders? Many morphological details reinforce the proto-lamprey impression:
But other characters reminiscent of gnathostomes or possibly the ancestral vertebrate condition:
Potential synapomorphies with Gnathostomata: (Therefore of Crown Vertebrata)

Euconodonta (Cambrian - Triassic)
According to the parsimony analysis of Miyashita et al., 2019, these are stem cyclostomes. Their Bayesian analysis placed them as stem hagfish. Either result is plausible.
Total Group Gnathostomata
Thelodonti (Ordovician - Devonian)

Morphology:
The hard skeleton of thelodonts consists entirely of odontodes - small scales that usually disarticulate when the animal dies. Odontodes are distinctive, consisting of dense and porous dentin layers around a pulp cavity, capped with enamel like a vertebrate tooth (!) Indeed, odontode-derived external scales and teeth that in most vertebrates line the jaws and pharyngeal elements are histologically and developmentally similar.
They form through inductive relationships between stratified ectodermal epithelium that secretes enamel and (depending on where on the body one is) out-pouching papillae of mesenchyme or mesenchyme-like neural crest that form dentin and pulp. Interestingly, although the pharynx is lined with endodermal epithelium, this can be co-opted to behave like enamel-forming ectoderm where endodermal cells are influenced by adjacent ectoderm that invades gill openings during development (Huysseune et al., 2021.)
Some thelodonts display the earliest known instances of odontodes invading the oral and pharyngeal cavities, even though they lack jaws. Rücklin et al., 2011 describe the odontodes of Loganellia scotica in detail.
Note: from this point on the tree onward, vertebrates generally retain odontodes or their derivatives, regardless of any other kind of skeletal ossification they may have.
Other thecodont features include:
- No proper paired fins, but many taxa have paired anterior triangular "fin folds" without skeleton or muscle that appear dorsal to the presumed position of the gill openings.
- One dorsal and one anal fin
- Hypocercal caudal fin.
- Paired olfactory capsules (in contrast to the vertebrates we have considered so far)
- Some well-preserved specimens appear to show a sediment-filled stomach, suggesting that they were deposit feeders.
- A series of eight - nine branchial (gill) openings
Furcacaudaformes: (Silurian - Devonian) While most thelodonts were slightly flattened dorso-ventrally, this group is morphologically distinctive in being:
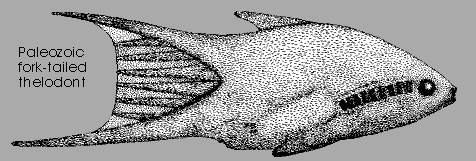
- laterally compressed
- distinct stomach present (it's earliest appearance in fossil vertebrates)
- Forked tail with major dorsal and ventral lobes, each of which supports up to four intermediate lobes.
- Paired fin folds present behind and below the gill openings. (In contrast to other thelodonts. Note: At least one taxon Shielia taiti has both types of fin folds. Are the two fin-fold types homologous?)
Thelodont monophyly: Is not assured. Recent analyses have (nearly) supported monophyletic Thelodonti:
- Donaghue and Smith, 2001 found non-furcacaudiform thelodonts to be monophyletic, with the furcacaudiforms forming a paraphyletic assemblage.
- Wilson and Märss, 2004 Found Thelodonti to be monophletic with Furcacaudiformes to be a basal clade.
Synapomorphies of Thelodonti and jawed-vertebrates:
- Dermal denticles with distinct root, crown, and pulp-cavity.
- Paired nasal capsules.

Pteraspidomorphi
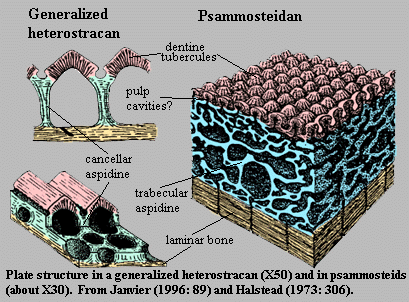
(Cambrian (assuming Anatolepis) minimally Ordovician- Devonian).The earliest well-preserved vertebrate, the Ordovician form Sacabambaspis, ironically represents a more derived form of hard tissue, in which individual denticles are integrated into broad head-shield composite elements and joined to one another through dermal layers of aspidin, a composite of thelodont-like denticles, dentin, and acellular bone. These shields are the first vertebrate elements that are bone-like in outward structure. Their possible functions include:
- Armor
- A reservoir of calcium and phosphate for use by the animal's metabolism.
- A solid skeleton against which muscles can act.
By the Silurian, many Sacabambaspis - like creatures are known, E.G. Astraspis (right).

Morphology:
-
- Two armor shields, one dorsal and one ventral.
- Armor plates covering each gill opening.
- The mouth is opened and closed by row of narrow parallel plates in the lower "lip."
- Heavy placoid scales on posterior half of body.
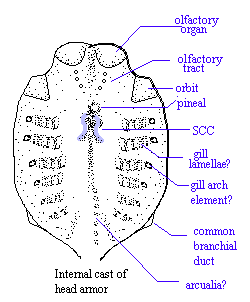
- The dorsal and ventral head shields carry impressions of internal structures including:
- Eyes
- Otic capsules with impressions of two sets of semicircular canals.
- Paired nasal capsules, but with no evidence of nostrils. The capsules might have communicated with a medial nasohypophyseal duct. (but see Galeaspida, below, for an alternative.)
- Branchial (gill) pouches with possible branchial arch elements external to the gills as in lampreys.
- Bony canals for lateral line system.
- Ossified cranial elements are known only from Eriptychius (Ordovician). They take the form of a set of separate preorbital elements unlike the separate cranial cartilages of lampreys or the consolidated neurocrania of gnathostomes (Dearden et al., 2023.)
- Notochord invades lower lobe of the hypocercal tail.
- There is no suggestion of dorsal, anal, or paired fins.
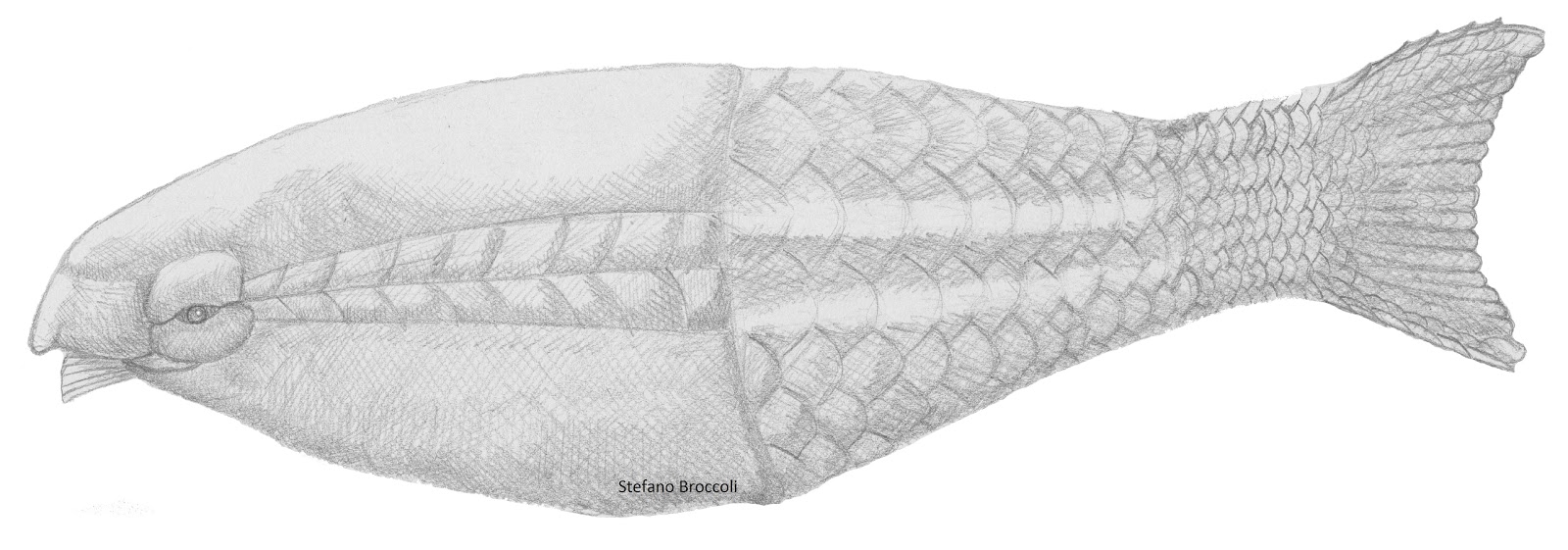
Heterostraci: The great majority of pteraspidomorph diversity consists of Heterostraci, such as Pteraspis right.

Their synapomorphy:
- All branchial openings are covered by single armor plates, so that there are only two excurrent gill openings.
Without a living analog, speculations about the ecology of these creatures is very limited. We note the following:

Feeding:
- If the distribution of pharyngeal slits is any indicator, they had a substantial pharynx.
- They lacked jaws, or even the keratinous structures that hagfish and lampreys use to process food. The implication was that they were suspension or deposit-feeding - filtering particulate matter. They may have lived like an armored version of ammocoetes (larval lampreys).
Janvier, 1997 suggests that the tail structure of basal pteraspidomorphs like Athenaegis (right) resembles that of furcacaudate thelodonts sufficiently to be a possible synapomorphy, suggesting that pteraspidomorphs might be derived from within Thelodonti. (According to Pradel et al., 2007, the tail of the early pteraspidomorph Sacabambaspis was strongly hypocercal, however.)
Synapomorphy of Pteraspidomorphi and jawed-vertebrates:
- Plate-like dermal bone formed in part from the coalescence of odontodes.

Issue: Transformation of feeding mechanics:
Remember the rod-and-pulley oral mechanism found in hagfish and lampreys, and inferred in euconodonts? Although the internal feeding apparatus of Jamoytius, Euphanerops, anaspids, thelodonts, and pteraspidomorphs is poorly know, so far, we have not surveyed any animal that could not possibly have had such a feeding mechanism:- those of pteraspidomorphs would seem to have required some mechanism to open the plates of the lower lip by pushing them forward
- rare well-preserved thelodonts display a mid-line structure (interpreted at right as the "esophagus") that could indicate a rod-shaped cartilage (among other possibilities). One could envision these working in concert with denticles lining the mouth and pharynx.
Issue: True Bone:
Before proceeding, a note: The cells that secrete and maintain hard tissue may be locked within it, yielding cellular bone. Seen in larger bony elements. Cellular bone forms in two ways:- Dermal bone: Laid down as a two-dimensional membrane. (E.g. human cranial bones). Ancestrally these formed near the body surface, but in derived vertebrates, their derivatives may invade deeper parts of the body.
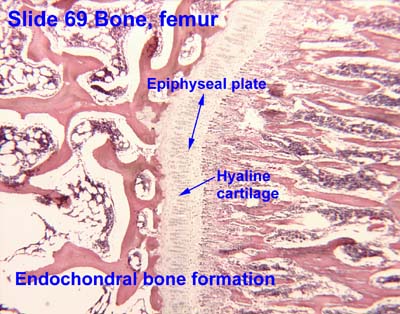
Endochondral bone cross-section from University of Oklahoma Health Sciences Center
Interactive Histology Atlas - Cartilage bone: Three-dimensional bone that is preformed in cartilage. (E.g. human limb bones). The cartilage, in turn, is preformed by condensations of mesenchyme cells (remember?), amoeboid mesodermal connective tissue of the embryo. Cartilage bone makes its evolutionary debut in the skeleton of the braincase but is widespread in vertebral columns and the skeleton of the limbs of derived vertebrates.
However cartilage bones in living vertebrates actually form in two ways:
- Perichondral ossification: Bone formation begins at the perichondrium - the membrane surrounding the cartilage precursor.
- Endochondral ossification: Bone formation begins in the interior of the cartilage precursor.
Note: Among living vertebrates, both types of ossification occur in cartilage bone, so neontologists and human anatomists call cartilage bone "endochondral bone." Paleontologists must distinguish them, however, because perichondral ossification evolved first and many fossil vertebrates have perichondrally ossified bone without the endochondral component.


Galeaspida:
Restricted to southern China and Indochina, then a separate continent. (Silurian - Devonian)
Morphology:
- large flat head shield enclosing a perichondral bone braincase. The head shield is a composite of the perichondral braincase and composite dermal bone like that of pteraspidomorphs, except that this is proper cellular dermal bone, not aspidin.
- Head shield encloses lateral-line system.
- large nasopharyngeal opening in upper front of head shield (presumably used for inspiration of respiratory water.) leads to:
- paired olfactory capsules
- hypophyseal duct
- mouth and pharynx.
- mouth is ventral
- the endocast (impression of the brain on the inner surfaces of the braincase) indicates the presence of a paired cerebellum. Note: hagfish and lampreys lack this, gnathostomes have it, and the fossils of pteraspidomorphs are ambiguous.
- Braincase encloses openings for sense organs:
- Orbits - the bony eye-sockets
- Pineal foramen - for the pineal eye.
- Pharynx is large with up to 45 pairs of gill openings, suggesting suspension feeding.
- The floor of the pharynx is made of distinct plates that could be moved to expand the pharyngeal cavity and draw in water. There is not evidence (or room) for a "rod-and-pulley" tongue mechanism.
- Caudal fin diphycercal or slightly hypocercal.
- Galeaspids are the first vertebrates in which the nasal capsules are separated from the hypophyseal duct. (Gai et al., 2011)
- The discovery of Tujiaaspis vividus shows us the postcranium in detail. (Gai et al., 2022) Tujiaaspis vividus has three dorsal fins and paired fin folds that extend the length of the body. Arguably the basis for the evolution of pelvic (and pectoral?) appendages.
Synapomorphy of Galeaspida and jawed-vertebrates:
- Cellular bone
- Perichondral bone in brain-case
- Nasal capsules separate from hypophyseal duct
- Paired cerebellum
- Paired fin-folds or their derivative.

Osteostraci
(Silurian - Devonian): Endemic to Euramerica. Resemble galeaspids in having large horseshoe-shaped head shields that include an perichondrally ossified brain-case, but with major differences:
Synapomorphies of Osteostraci:
- Head shield contains three large sensory fields (two laterally and one on the midline). Internal channels link these to the otic capsules.
- An odd ventral fin flap laps beneath the caudal fin. This might be a modified anal fin.
- The floor of the pharyngeal chamber is flexible, facilitating inspiration of water.
- The oral and pharyngeal cavities seem to be separated by a velum.
- Posterior corners of the head can be drawn into cornual processes or "horns."
- Sclerotic ossicles support the sclera of the eyeball.
- Endolymphatic ducts connect the fluid filled labyrinth of the inner ear with the external environment. This is also observed in placoderm-grade gnathostomes and living Chondrichthyes.
- Notochord invades upper lobe of tail yielding heterocercal tail.
- One or two dorsal fins present
- Branchial openings slit-shaped. (Compare with lamprey)
- But the big thing: Paired pectoral fins (complete with calcified cartilage skeleton elements) present.
- Single median nasohypophyseal opening leads to a single olfactory capsule (like in lampreys) and to the hypophysis, and doesn't communicate with the pharynx - like the arrangement in lampreys.
Osteostraci are both informative and misleading. Their well-ossified brain-cases enable us to know their anatomy as in no other fossil jawless vertebrate, but aspects of this anatomy are enigmatic:

- What are the three "head fields?" Electric organs, elaborations of the lateral-line system, and outgrowths of the otic system are all suggested.
- Is the lamprey-like nasohypophyseal arrangement ancestral or secondarily derived? For decades, systematists linked osteostracans to lampreys because of it. Recently, the single nasal olfactory capsule and midline nasohypophyseal duct are considered to be a secondary reversal from a condition like that in galeaspids.
- Osteostraci share mesodentin with Katoporida, a thelodont taxon. Synapomorphy?
Synapomorphies of Osteostraci and jawed-vertebrates In any case, Osteostraci shares conspicuous synapomorphies with Gnathostomata:
- Endolymphatic ducts communicate with the exterior.
- Paired pectoral appendages with internal skeleton and muscle
- Heterocercal tail
- Two dorsal fins
- Sclerotic ossicles
- Slit-shaped branchial openings

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Pituriaspida:
(Devonian) A poorly known group with two known genera: Pituriaspis and Neeyambaspis from Queensland. Known only from their well-integrated, solid head shields. Generally similar to Osteostraci. Characterized by:- An elongate rostrum (snout)
- Nasohypophyseal opening beneath shield in front of mouth.
- Mouth and pharynx organized as in Osteostraci
- Paired openings of unknown function beneath orbits (eye sockets)
- Opening for pectoral fins present in head shield
- Head shield extends far posteriorly. Possibly (?) forming support for pelvic appendage (???)
More and better fossils, please.
Coping with history: You will surely encounter the obsolete term "ostracoderms" in the literature. This refers to the vast paraphyletic grade-group of armored jawless vertebrates whose diversity we have surveyed.
Major Outstanding Issues:
- The position of soft-bodied craniates like Myllokunmingia and Metaspriggina
- The origin and homology of paired fins
- Thelodonts - monophyletic or paraphyletic?
- The evolution of the organization of the head (braincase, nasohypophyseal duct, nasal capsules, etc.) in vertebrates lacking cartilage bone.
- The evolution of the weird nasohypophyseal duct of Osteostraci.
- Raisa Bailon-Zambrano et al.. 2024, The sclerotome is the source of the dorsal and anal fin skeleton and its expansion is required for median fin development. Development, 151(24), 1-14.
- Derek Briggs, Euan Clarkson, and Richard Aldridge. 1983, The Conodont Animal. Lethaea, 16(1), 1-14.
- Simon Conway Morris and Jean-Bernard Caron. 2014. A primitive fish from the Cambrian of North America. 512, 419-422.
- Mason Dean and Ron Shahar, 2012. The structure-mechanics relationship and the response to load of the acellular bone of neoteleost fish: A review. Journal of Applied Ichthyology, Feb.
- Richard P. Dearden, Agnese Lanzetti, Sam Giles, Zerina Johanson, Andy S. Jones, Stephan Lautenschlager, Emma Randle, and Ivan J. Sansom 2023. The oldest three-dimensionally preserved vertebrate neurocranium. Nature 621, pages782–787.
- Philip Donoghue and M Paul Smith, 2001.The anatomy of Turinia pagei (Powrie), and the phylogenetic status of the Thelodonti. Transactions of the Royal Society Edinburgh, Earth Sciences. 92: 15-37.
- Philip C. J. Donoghue and Joseph N. Keating. 2014. Early vertebrate evolution. Palaeontology. Early View online version August 16, 2014.
- Todd Feinberg and Jon Mallatt. 2013. The evolutionary and genetic origins of consciousness in the Cambrian Period over 500 million years ago. Frontiers in Psychology 4(667):667
- S. E. Gabbott, R. J. Aldridge, J. N. Theron. 1995. A giant conodont with preserved muscle tissue from the Upper Ordovician of South Africa. Nature 374, pages800–803.
- Zhikun Gai, Philip Donogue, Min Zhu, Philippe Janvier, and Marco Stampanoni. 2011, Fossil jawless fish from China foreshadows jawed vertebrate anatomy. Nature, 476, 324-327.
- Zhikun Gai, Qiang Li, Humberto G. Ferrón, Joseph N. Keating, Junqing Wang, Philip C. J. Donoghue & Min Zhu. 2022, Galeaspid anatomy and the origin of vertebrate paired appendages. Nature, 609, 959–963.
- Nicolas Goudemand, Machael Orchard, Severine Urdy, Hugo Bucher, and Paul Tafforeau. 2011, Synchrotron-aided reconstruction of the conodont feeding apparatus and implications for the mouth of the first vertebrates. Proceedings of the National Academy of Sciences, 108(21), 8720-8724.
- Yara Haridy, Markus Osenberg, Andre Hilger, Ingo Manke, Donald Davesne, and Florian Witzmann. 2021. Bone metabolism and evolutionary origin of osteocytes: Novel application of FIB-SEM tomography. Science Advances 7(14).
- Guillaume Houée, Nicolas Goudemand, Damien Germain, Jérémie Bardin. 2024. Paleo-evo-devo implications of a revised conceptualization of enameloids and enamels. Biological Reviews December 2024.
- Ann Huysseune, Robert Cerny, P. Eckhard Witten. 2021. The conundrum of pharyngeal teeth origin: the role of germ layers, pouches, and gill slits. Biological Reviews 97(1): 414-447.
- Philippe Janvier. 1993. Patterns of diversity in the skull of jawless fishes. In The Skull (ed. J. Hanken and B. K. Hall), Vol. 2, pp. 131 - 188. The University of Chicago Press.
- Philippe Janvier. 1997. Thelodonti. In The Tree of Life web site, http://tolweb.org/tree?group=Thelodonti&contgroup=Vertebrata, recovered 12/29/2015.
- Philippe Janvier. 2006. Lamprey-like gills in a gnathostome-related Devonian jawless vertebrate. Nature 440, 1183-1185.
- Philippe Janvier. 2007. Homologies and Evolutionary Transitions in Early Vertebrate History. In Major Transitions in Vertebrate Evolution. (eds. Hans-Dieter Sues and Jason Anderson), pp. 57 - 121 Indiana University Press, Bloomington.
- Philippe Janvier. 2007. The anatomy of Euphanerops longaevus Woodward, 1900, an anaspid-like jawless vertebrate from the Upper Devonian of Miguasha, Quebec, Canada. Geodiversitas, 29(1):143-216
- Rudy Lerosey-Aubril and Javier Ortega-Hernandez. 2024. A long-headed Cambrian soft-bodied vertebrate from the American Great Basin region. Royal Society Open Science 11(7):240350.
- Tiiu Märss and A Ritchie (1998), Articulated thelodonts (Agnatha) of Scotland, Transactions of the Royal Society Edinburgh, Earth Sciences. 88: 143-195.
- Tetsuto Miyashita, Michael I. Coates, Robert Farrar, Peter Larson, Phillip L. Manning, Roy A. Wogelius, Nicholas P. Edwards, Jennifer Anné, Uwe Bergmann, A. Richard Palmer, and Philip J. Currie. 2019. Hagfish from the Cretaceous Tethys Sea and a reconciliation of the morphological–molecular conflict in early vertebrate phylogeny. Proceedings of the National Academy of Science published ahead of print January 22, 2019.
- Takayuki Onai. 2023. Organization of the body wall in lampreys informs the evolution of the vertebrate paired appendages. Journal of Morphology 284:3 e21559.
- Pander, Heinz. 1856. Monographie der Fossilen Fische des silurischen Systems der Russisch-Baltischen Gouvernements (Monograph of fossil fish from the Silurian stratum of the Baltic regions), St. Petersburg
- Jane C. Reeves, Roy A. Wogelius, Joseph N. Keating, Robert S. Sansom. 2023. Lasanius, an exceptionally preserved Silurian jawless fish from Scotland. Palaeontology 66(2).
- Alan Pradel, Ivan. J Sansom, Pierre-Yves Gagnier, Ricardo Cespedes, Philippe Janvier. 2007. The tail of the Ordovician fish Sacabambaspis. Biology Letters 3(1).
- Martin Rücklin, Sam Giles, Philippe Janvier, and Philip Donoghue. 2011. Teeth before jaws? Comparative analysis of the structure and development of the external and internal scales in the extinct jawless vertebrate Loganellia scotica. Evolution and Development 13(6) 523-532.
- Robert Sansom, Kim Freedman, Sarah Gabbott, Richard Aldridge, and Mark Purnell. 2010. Taphonomy and affinity of an enigmatic Silurian Vertebrate Jamoytius kirkwoodi White. Palaeontology 53(6), 1393-1409.
- D-G. Shu, H-L. Luo, S. Conway Morris, X-L. Zhang, S-X. Hu, L. Chen, J. Han1, M. Zhu, Y. Li, and L-Z. Chen. 1999. Lower Cambrian vertebrates from south China. Nature 402, 42-46.
- Qingyi Tian, Fangchen Zhao, Han Zeng, Maoyan Zhu, and Baoyu Jiang. 2022.
Ultrastructure reveals ancestral vertebrate pharyngeal skeleton in yunnanozoans. Science, 377(6602), 218 - 222.- Mark Wilson and Tiiu Märss. 2009. Thelodont phylogeny revisited, with inclusion of key scale-based taxa. Estonian Journal of Earth Sciences. 58, 297-310.
- Derek Briggs, Euan Clarkson, and Richard Aldridge. 1983, The Conodont Animal. Lethaea, 16(1), 1-14.