
Vertebrata within Metazoa

Metazoa:
Animals in the broadest sense.The phylogeny of major animal groups appears at right. Included are:
- Porifera The sponges. (Quotation marks indicate that the group is probably paraphyletic
- Ctenophora The comb-jellies.
- Cnidaria Corals, anemones, jellyfish, etc.
- Spiralia Mollusks, brachiopods, bryozoans and kin.
- Ecdysozoa Arthropods and relatives.
- Deuterostomia The group containing vertebrates and echinoderms.
- Porifera
- Ctenophora
- Cnidaria
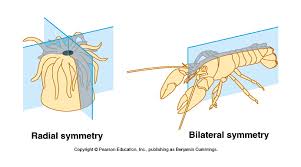
- proper bilateral symmetry
- flow-through guts with distinct mouths and anuses
Bilateria:
Animals with:- Bilateral symmetry
- A flow-through gut in which we can distinguish a mouth and anus.
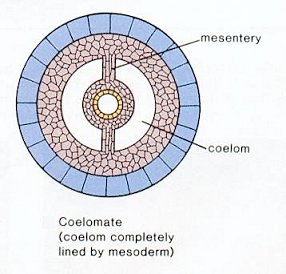
- Mesoderm: A third tissue layer occupying the space between endoderm and ectoderm, giving rise to many internal structures, including...
- The coelom, or body cavity, made many types of life strategies possible because its possessors could truly move in a purposeful manner.
- Among such animals, we see two major branches identified by distinct patterns of embryology. (Bilaterian developmental issues are covered in a later lecture.)
- Protostomia
- Deuterostomia
Protostomia contains the vast majority of animal diversity, including many familiar animals. Among them are two major groups:
- Ecdysozoa
- Spiralia.

- Ecdysozoa includes Arthropods among others.

- Spiralia includes, among others mollusks and annelids.
There is a trend in many bilaterian groups including active animals toward the evolution of special sense organs (eyes, antennae) and toward concentrating them at the front end along with an expansion of the nervous system to deal with the information they provide (heads).
And yet, these structures originate independently. Consider some close relatives of the major groups: Priapulids for ecdysozoans, and phoronids for Spiralians.
Deuterostomia:
Living deuterostomes were first identified by "deuterostomous" developmental characters (later). This identification had little to do with their outward morphology. Indeed, the major groups are an odd assortment:

Cyprinius carpio
- Cephalochordata - Amphioxus
- Urochordata - tunicates
- "Craniata" - vertebrates and their closest relatives (right)

Leopard sea cucumber Bohadschia argus
- Crinoidea - sea lilies
- Asteroidea - starfish
- Echinoidea - sea urchins
- Ophiuroidea - brittle-stars
- Holothuria - sea cucumbers (right)
Hemichordata: (C. - Rec.) Including

- Enteropneusta - acorn worms (right)
- Pterobranchia - pterobranchs and graptolites
Yet wonderfully, the illumination of Cambrian fossils on the stems of each of these groups has provided significant, if imperfect resolution. Of particular importance:
Vetulicolia: Known from the classic fossil localities of Chengjiang (Yunnan Province, China) and Sirius Passet (Greenland). These animals encompass a range of shapes but have these characteristics in common:

- Lacking hard parts, but with an outer skeleton of plates of cuticle.
- Body divided into a barrel-shaped front portion and a tail with seven distinct segments.
- At the front is a large mouth without feeding appendages. (Indeed, their are no appendages at all.)
- The front portion encloses a large open space
- Five pairs of openings penetrate the sides of the front segment along a pair of grooves.
- The tail is a laterally compressed and moved from side to side.
The consensus interpretation (Vinther et al., 2011) is that:
- The front segment represents a pharynx - a large chamber through which particulate-laden water is filtered for feeding.
- The tail is homologous to the tails of chordates
- Vetulicolians (along with a few other oddballs) branch from the base of Deuterostomia, although some researchers consider them to be closer to the crown of Chordata. (See Garcia-Bellido, et al., 2014.)
How did deuterostomes (and bilaterians) originate? Han et al. 2017 report on Saccorhytus coronarius, a millimeter-scale critter from the basal Cambrian with a mouth and lateral openings but no anus. their phylogenetic analysis places it as a basal vetulicolian. Maybe. This points to the possibility that deuterostomes (and other bilaterians) originated as part of the minute meiofauna of the earliest Cambrian.

Deuterostomia:
(Cambrian - Rec.) Vertebrate paleontologists have long predicted that the ancestral vertebrate would be, in essence, a swimming pharynx. The interesting surprise is that this seems to describe deuterostomes in general. This information enables us to propose morphological synapomorphies of Deuterostomia:- Deuterostomous development
- A pharynx with phayrngeal openings
- A segmented tail that undulates laterally
Deuterostome phylogeny is characterized by two major groups:
- Ambulacraria: Echinodermata and Hemichordata
- Chordata: Vertebrates and their relatives.


Ambulacraria:
Recent molecular phylogenies indicate sister taxon relationship between Echinodermata and Hemichordata. Most members are suspension or deposit feeders, although some obtain food in more interesting ways.Potential synapomorphies:
- Molecular characters
- Feeding appendages (proboscis and feeding appendages of hemicordates, ambulacra of echinoderms) containing coelomic extensions
- Similar larval forms (right)

Hemichordata:
(Cam - Rec.) Solitary or colonial suspension and deposit feeders who use ciliated appendages to concentrate particle-rich water in pharynx, where it is filtered.Characteristics:
- Bodies divided into proboscis, collar, and trunk, each of which is invested with its own part of the three-part coelom (right):
- Cilia on the proboscis move food particles to mouth.
- Collar encircles mouth and proboscis.
- Trunk contains a large pharynx with pharyngeal openings (homologous to gill slits)
- Open circulatory system present, with blood propelled by contractions of primary blood vessels. No hemoglobin.
- Cilia on the proboscis move food particles to mouth.
- Dorsal nerve cord, hollow in some parts of collar.
Diversity:
Enteropneusta: "Acorn worms": (Middle Cambrian - Quaternary)
- Largish (5 cm - >2m) worm-like burrowing suspension or deposit feeders.
- Pharynxes large and perforated by many pairs of gill openings.
- Acorn-shaped proboscis gives common name
- Invariably solitary.
Pterobranchia: (Cambrian - Quaternary)

Characteristics:
- Tiny (<2 mm.) colonial critters that secrete branching colony consisting of proteinaceous tubes.
- The proboscis is developed into a cephalic shield, used to secrete the proteinaceous material of the colony structure.
- The collar is developed into one to five pairs of ciliated particulate food gathering arms that sport cilia-covered tentacles that transport food particles to mouth.
- Only one pair of pharyngeal openings.
- Invariably colonial.
- During the last twenty years, it has become evident that graptolites - common fossils from the early Paleozoic, are pterobranch colonies.
 Yunnanozoans: A final fossil group from Chengjiang (Early Cambrian), these creatures (including Yunnanozoon (right) and Haikouella) were described in the 1990s. These animals definitely had:
Yunnanozoans: A final fossil group from Chengjiang (Early Cambrian), these creatures (including Yunnanozoon (right) and Haikouella) were described in the 1990s. These animals definitely had:
- An integument (outer layer) of cuticle.
- Trunk muscle divided into blocks (myomeres)
- Seven pharyngeal openings with possible gill filaments.
- In some interpretations, both ventral and dorsal nerve cords.
- Brains
- Special sense organs (eyes, otic capsules, etc.)
- Chevron-shaped myomeres.
- stem chordates (Mallatt and Chen, 2003; and others)
- stem ambulacrarians (Shu, 2003)
- stem hemichordates (Shu et al., 1996)
- cephalochordates (Chen et al., 1995)
- stem deuterostomes (Shu et al. 2001)
- stem bilaterians (!) (Dewel, 2000)
- As of 2022, as stem vertebrates (!) (Tian, et al., 2022) based on details of their pharyngeal bars. (An issue for another lecture.)
Echinodermata:

Shingle urchin - Colobocentrotus atratus
- Exclusively marine: Echinoderms lack osmoregulatory mechanisms that might allow them to live in brackish or fresh water.
- Skeleton is internal test comprised of individual plates of porous high-Mg calcite. In life, the pores are occupied by a protein matrix and dermal cells. Such skeletal tissue is known as stereom. Carbonate petrologists typically call the pores "meat holes." individual birefringent elements
- One portion of the coelom develops into water vascular system. (Dissection)
- Living (and most but not all) fossil forms have strict five-part symmetry.
And yet, the most basal Cambrian forms approach bilateral symmetry and have been shown by Rahman et al., 2015, probably to possess a pharyngeal filtration system.
Chordata:
The sister taxon of Ambulacraria. Contains:- Urochordata: Tunicates (aka "sea-squirts")
- Cephalochordata: Living Branchiostoma (aka amphioxus, aka lancelets. Deal with it.)
- Vertebrata: Creatures with heads, special sense organs, vertebral columns.

Schematic of ancestral vertebrate (showing chordate characters plus some)
from Radinsky 1987. The Evolution of vertebrate Design.
- Body stiffened by a notochord - a rod made of cartilage-like tissue and fluid wrapped in a tight membrane - at some point in life
- Dorsal nerve cord hollow for entire length.
- Mucus-secreting endostyle at ventral midline of pharynx.
- Post-anal tail.
- Body musculature divided into chevron-shaped myomeres. (Myomeres - segmented blocks of trunk muscle are also present in vetulicolians and yunnanozoans, but in them are not chevron-shaped.
Pikaia - Almost a chordate:

The Burgess Shale fossil Pikaia gracilens, known from hundreds of specimens, is often cited in popular literature as the "earliest chordate." In fact:
- It is not the earliest. That would be either Myllokunmyngia, the similar Haikouicthys, or Cathaymyrus from Chengjiang.
- The most recent redescription by Mussini et al., 2024, shows that it is not quite a chordate as it lacks a post-anal tail or endostyle, and its notochord can't reliably be identified. Link to interpretation.
- Significantly, it does seem to have a tiny head, with a pair of antennae.
- Chevron-shaped myomeres

Cephalochordata:
(Cambrian - Quaternary) Represented by the living Branchiostoma.Nomenclature: Earlier names have priority in scientific nomenclature. Unfortunately, the publication in which this animal was originally named (Branchiostoma) was largely ignored, and it was named a second time (Amphioxus). Additionally, there had been a common name (Lancelet). When the dust settled, Branchiostoma was acknowledged the proper scientific name, and amphioxus and lancelet are both considered common names. Live with it.
Anatomy: In many ways, Branchiostoma is like a modified version of Pikaia. Similarities include:
- Food is captured by filtration of water through a pharynx
- Muscles enabling the animal to swim by lateral undulations are divided in to chevron-shaped myomeres.
- Dorsal nerve cord.
- Mid-line dorsal and ventral fins are present forming a tail.
- Body supported by a robust notochord that extends from the tip of the snout to the end of the tail. This supports...
- The dorsal nerve cord is hollow for its entire length. There is no obvious brain, but anteriorly the nerve cord expresses genes found in the vertebrate forebrain and midbrain, and displays distinct anterior and posterior cerebral vesicles. Gene expression suggests that their junction corresponds to the vertebrate hypophysis (Lacalli, 2024. (Figure C in linked image.)
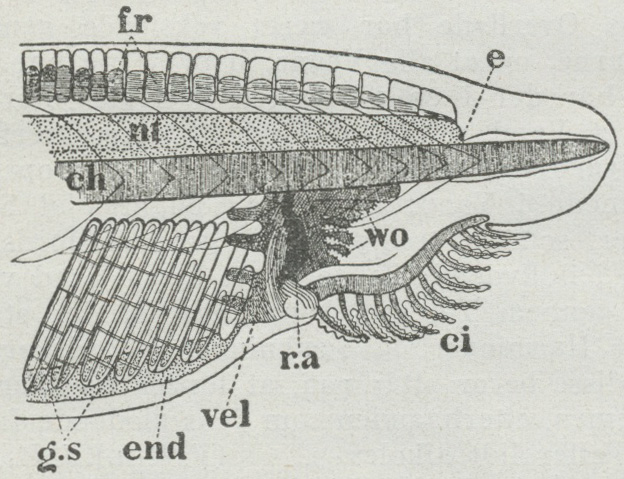
- A distinct oral chamber is separated from the pharynx by a velum
- the pharynx is:
- Large
- Pharyngeal openings are tall slits separated by pharyngeal bars.
- Water filtered through pharyngeal slits by ciliary action enters an atrium - an outer chamber that envelops the pharynx, before it is expelled.
- A gland called the endostyle occupies the ventral mid-line of the pharynx. It secretes mucus that is carried along the pharyngeal bars by cilia. This mucus, along with anything digestible that sticks to it, is transported to the esophagus and gut tube.
- The post-anal tail tail extends well past the anus.

Branchiostoma life position
Anatomical details:
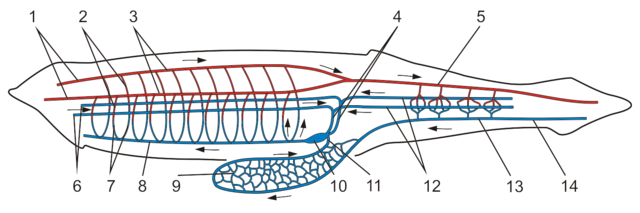
- Although there is no heart, the circulatory system is closed. Blood flows through a ventral aorta to the branchial bars, up through them, and into a dorsal aorta through which it is taken to the body.
- The dorsal nerve cord is hollow and there is a slight anterior enlargement. (Indeed, according to Schubert et al., 2006 genes expressed in vertebrate brains are also expressed in this region, providing some basis for the establishment of homology.
- Hatschek's pit, a chemosensory organ in the roof of the oral chamber, is homologous with the vertebrate hypophysis (pituitary gland) and marks the junction of the anterior and posterior cerebral vesicles.
- A diverticulum of the gut occupies the place of the liver in craniates.

Cathaymyrus diadexus from Dr. Ken Hooper
Virtual Natural History Museum Ottawa-Carleton Geoscience Centre
- Notochord extends to the front end of the body.
Cephalochordate fossil record: Sparse, but present as early as the Chengjiang lagerstätte in the form of Cathaymyrus diadexus (right).
Interpreting cephalochordates: For decades, cephalochordates have been presumed to be closely related to Chordata. They are so close morphologically. The Wizard need only give them hearts and brains. The next group, in contrast, seems more distant.

Urochordata:
(Sparse record Cam. - Rec.) Also known as "tunicates" or "sea squirts." As adults, the body is dominated by a huge pharynx used for suspension-feeding. Adults of readily accessible species are typically attached to some hard surface, however many are free-swimming. They may be solitary or colonial.Anatomy:
- Their bodies consist of a large basket-like pharynx perforated by numerous slits. On one side, the endostyle secretes mucus which is transported across the inner surface of the pharynx by cilia, capturing food particles as it goes.
- Entire body enclosed by a "tunic" made primarily of tunicin, a unique complex sugar. Water enters pharynx through a "mouth" and, after filtration, enters an atrium from which it is expelled through a separate opening, the atriopore.
- The anus also opens into the atrium,
- Heart and open circulatory system present. Blood flow reverses periodically.

- a notochord
- a hollow dorsal nerve cord
- muscles arranged into myomeres

When your instructors completed their PhDs, the consensus of deuterostome phylogenies held that:
- Hemichordata was the sister taxon to Chordata. Synapomorphy: Pharyngeal openings.
- Cephalochordata was the sister taxon of Vertebrata.
- The Burgess shale fossil Pikaia was a cephalochordate because it resembled Branchiostoma and had no distinct head.
- Urochordates we considered to represent an ancestral form, from which cephalochordates and craniates might have evolved by pedomorphosis.
Discoveries that have changed that view:
- Vetulicolians and Yunnanozoans provide a picture of the ancestral deuterostome as a "swimming pharynx." Thus:
- The swimming morphology and segmentation of craniates, cephalochordates, larval urochordates, and Pikaia are plesiomorphies.
- Same for the presence of pharyngeal openings in hemichordates and chordates.
- If Larvacea represents the basal condition for Urochordata (Swalla et al. 2000) then urochordates also arose from "swimming phayrnges." (Indeed, Garcia-Bellido et al. 2014 propose that vetulicolians are actually stem urochordates. We await confirmation.)
- Strong molecular evidence for Olfactores and Ambulacraria that doesn't imply improbable reversals and convergences.
- Re-description and "demotion" of Pikaia.
Relationships within Chordata:
Olfactores:
Synapomorphies of craniates and urochordates:- Neural crest sells: Ectodermal cells that slough off of the forming neural tube that migrate to various regions of the body, giving rise to a range of anatomical features. Although neural crest homologs (marked by gene expression) may be present in Branchiostoma Holland et al., 1996, morphologically identifiable neural crest is absent, as are ectodermal placodes.
- In urochordates, migratory neural crest cells exist, but only control the distribution of pigment in the skin. Although there are no special sense organs, the atrium arises through the invagination of an ectodermal placode.

Vertebrata
(Cambrian - Recent). Last common ancestor of lampreys and gnathostomes and all descendants. Pretty much includes all chordates with heads. Unambiguous synapomorphies include:- Head including:
- Anterior expansion of nerve cord into brain.
- Special sense organs:
- Olfactory capsules for olfaction
- Eyes for vision:
- large paired lateral eyes for image forming
- simple, mid-line pineal eye as light-meter
- Otic capsules for hearing
- A hypophysis (AKA pituitary gland) occupies a recess in the mid-line floor of the brain-case. It is a composite organ containing both endocrine and brain tissue. Ancestrally, it communicates with the exterior via a nasohypophyseal duct.
- Neural crest tissue forms significant portions of the head, gill-arch skeleton, peripheral nervous system, and pigmentation.
- Water pumped through pharynx by muscles instead of my ciliary action.
- The bars separating the slits of the pharynx support special gas-exchange organs - gills.
- Larval endostyle transforms into adult thyroid gland
In the post-cranium:
- Arcualia - cartilaginous precursors to vertebrae. These are long known to extend the entire length of the skeleton in lampreys but simple versions were described by Ota et al., 2011 in the caudal (tail) skeleton of hagfish.

Hagfish arcualia - W - shaped myomeres - an elaboration of the simple chevrons of Chordata.
- Caudal fin stregthened by cartiligenous radials
- In the gut:
- Liver and pancreas present (formed through endoderm-mesoderm induction)
- Digestive system invested with smooth muscular lining (rather than cilia)
- Circulatory System
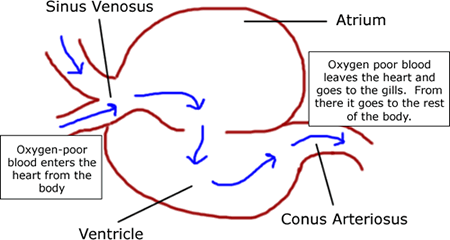
- Two chambered heart
- Hemoglobin for oxygen transport
- Erythrocytes (red blood cells) to contain hemoglobin
- Increased size (an order of magnitude)
- Cartilaginous endoskeleton: including fin rays and brachial arches
- Orders of magnitude bigger than any non-craniate chordate
- Much more active than non-craniate chordates.
In terms of metabolic rate and aerobic capacity, only cephalopod mollusks and some arthropods are comparable to vertebrates.
Vertebrate diversity:
First, an inventory of the living groups:
- Hyperotreti - the Myxinoidea (hagfish) and relatives
- Hyperoartia - the Petromyzontiformes (lampreys) and relatives
- Gnathostomata - the jawed vertebrates

Hyperotreti and Myxinoidea:
(Hagfish - Carboniferous - Quaternary). Synapomorphies of Hyperotreti:- Comb-shaped keratinous biting mouthparts
- Four pairs of tentacles surround the mouth.
- Impressive mucus glands
Other characteristics:
- The skeleton consists of the notochord and specialized cartilages of mouth. The latter take the form of a mid-line rod-and-pulley upon which the tooth-plates are protracted and retracted.
- Two-chambers of the heart are poorly consolidated.
- Single nasohypophyseal opening leads to a nasohypophyseal duct opening onto:
- a mid-line olfactory capsule whose skeleton resembles a basket.
- the hypophysis
- and communicates posteriorly with the pharynx.
- Otic (inner ear) capsules have only one semicircular canal (as opposed to three in jawed vertebrates). But note: The hagfish semicircular canal has two distinct populations of hair cells, hinting at a complex evolutionary history.
- Five to fifteen pairs of gill openings.
But hagfish lack many features we see in other craniates:
- Any suggestion of arcualia outside of the tail. (In hagfish, the only arcualia form in the position of the hemal arches of more derived vertebrates.)
- A cartilage brain-case as part of their skeleton. Theirs is largely membranous.
- Extrinsic eye muscles: Hagfish eyes are simple and cannot be rotated in the head.
- Taste buds
- Electroreceptors in skin.
Not surprisingly, hagfish have almost no fossil record. Potential fossil hagfish include (in order of descending quality of fossil) the stem hyperotretans:

- Myxinikela (Carboniferous) from Mazon Creek. Distinct tentacles and nasal basket on a rather plump body. (Interpretation of head.)
- Myxineides (Carboniferous) an eel-shaped smear with evidence of hagfish-like tooth plates.
- Gilpichthyes (Carboniferous) another eel-shaped smear (but according to Miyashita et al., 2019, this is a stem lamprey!)
Within the crown of Myxinoidea, we have Tethymyxine (Cretaceous) from Miyashita et al., 2019, which displays all characteristic hagfish features.

Hyperoartia and Petromyzontiformes:
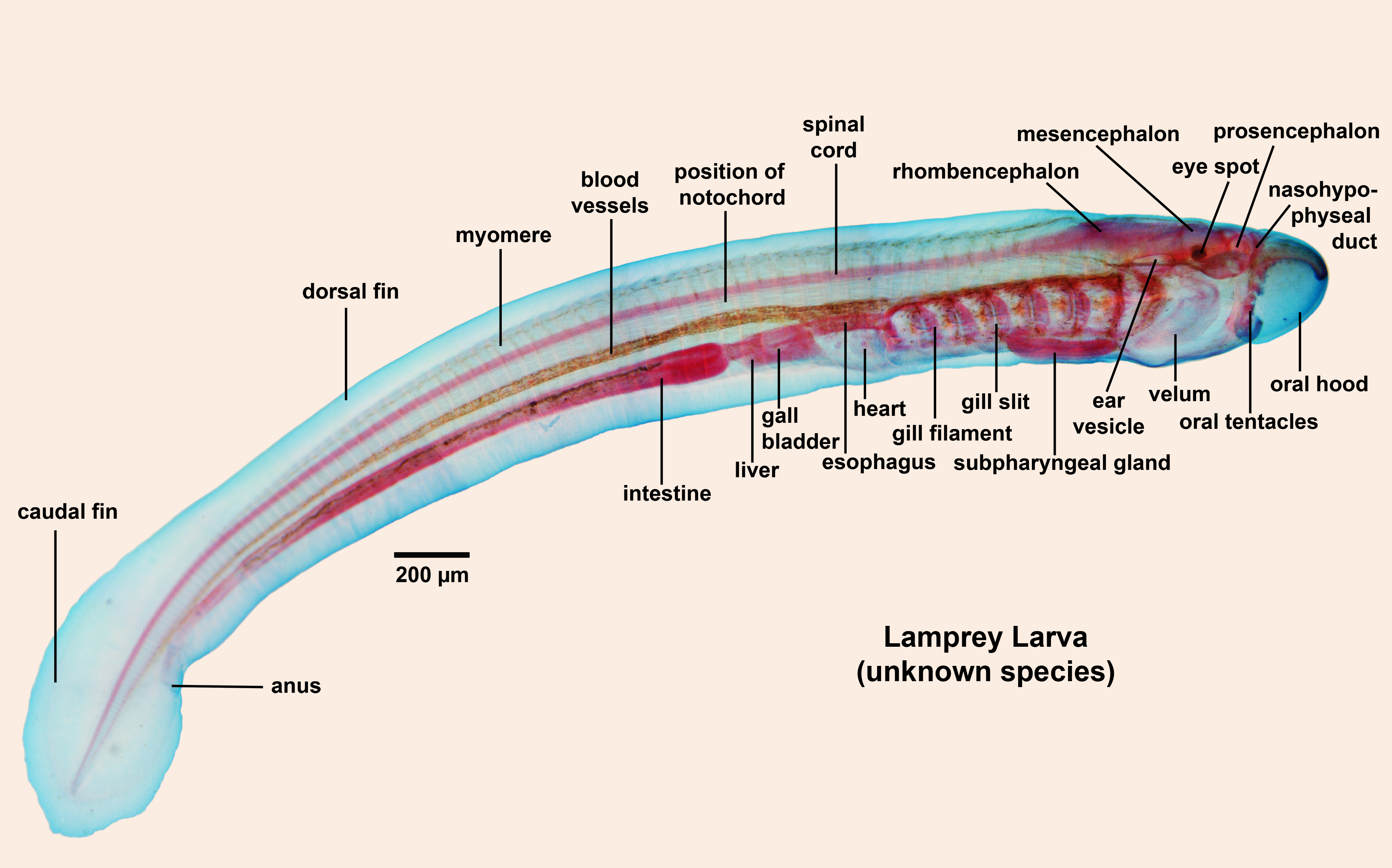
(Lampreys) - (Devonian - Recent): Characterized by:- Undergoes metamorphosis from suspension-feeding ammocoetes larva that looks like a brainy version of Branchiostoma to a parasitic adult.
- In adult, annular cartilage supports a large sucker surrounding the mouth armed with keratin "teeth". This is supported by other unique cartilage elements.
- Piston cartilage supporting a protrusable "tongue" armed with more keratinous denticles. Although distinct, this is also a rod-and-pulley arrangement similar to that of hagfish.
- Like hagfish in having a median nasohypophyseal duct that communicates with median nasal capsule and the hypophysis. Unlike hagfish (and like jawed vertebrates) the nasal duct does not lead to the pharynx. (The single nostril does still communicate with both the nasal capsule and hypophysis.)
- Branchial skeleton: the lamprey gill pouches are supported by a branchial basket whose cartilagenous elements are external to the gills.
- Seven pairs of gill openings.
- Tail bends downward slightly (barely hypocercal)
- Living taxa have two distinct dorsal fins, although fossils don't.
- Two vertical semicircular canals in the otic capsule. (In this and in other respects considered below, lampreys appear to have much in common with the final craniate group.)

Priscomayon riniensis (Late Devonian)
- Priscomyzon riniensis (Devonian - right) (Gess et al., 2006). Tiny at 4.2 cm.
- Hardistiella montanensis (Early Carboniferous) - Very small and lacking two distinct dorsal fins, but distinctly a metamorphosed adult.
- Mayomyzon pieckoensis (Late Carboniferous) - like Hardistiella but clearly showing annular and piston cartilages.
Now it gets strange: Because of vertebrates' close relationship with urochordates and cephalochordates, we have assumed that lampreys' suspension-feeding ammocoetes larval stage was an instance of ontogeny recpaitulating phylogeny. According to Miyashita, 2018, a solid growth series for Priscomyzon shows that it lacks the ammocoetes phase, and hatches out as a tiny parasite. Apparently the ammocoetes ontogenetic stage is a derived feature of modern lampreys! This challenges our basic assumptions about the ancestral vertebrate being a vetulicolian-ike suspension feeder. Could the common ancestor of craniates have, instead, been a deposit-feeder? But there is more lamprey-feeding confusion...

Tullymonstrum gregarium (Late Carboniferous)

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
.jpg){kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Gnathostomata:
(Silurian - Quaternary) The last common ancestor of jawed vertebrates and all of its descendants. Living gnathostomes are distinct from living either Hyperotreti or Hyperoartia in many respects. Conspicuously, they have- Jaws
- Branchial skeleton internal to gills, each set consisting of a series of hinged bars.
- Paired limbs: Distinct pectoral and pelvic fins.
- Three pairs of semicircular canals in otic capsules
- Reorganization of the mouth, pharynx, nasal capsules, and nasohypophyseal duct so that nasal capsules communicate directly to the exterior via nostrils.
The lifestyles of vertebrates were revolutionized by the ability to process large food items.

Relationships
Illuminating the relationship between Hyperotreti, Hyperoartia, and Gnathostomata is difficult and contentious. Two major hypotheses predominate that take their names from the positions they imply for lampreys:
- The Cyclostome hypothesis, in which Hyperotreti and Hyperoartia form a clade - Cyclostomi.
- The Vertebrata hypothesis, in which Hyperoartia group with Gnathostomata to make a monophyletic Vertebrata that excludes hagfish.
Cyclostomi:
(Devonian - Quaternary) Support for a close relationship of lampreys and hagfish is mostly molecular (E.G. Kuraku et al., 2009), however this support is widely replicated and very strong. Clear morphological synapomorphies are not obvious, but possible candidates include:- The rod-and-pulley arrangement of the tongue (Yalden, 2008), however as we will see in fossil jawless vertebrates, this might be plesiomorphic.
- Keratinous tooth-plates.
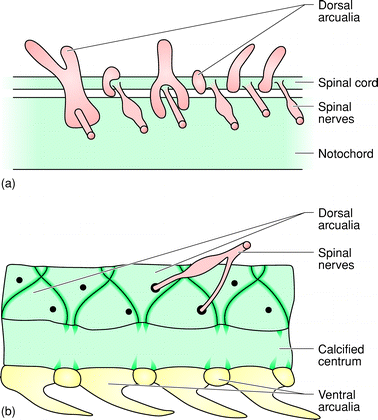
Evolutionary stages of chondrification of vertebrae:
Above - lamprey; below - shark
Vertabrata:
(Cambrian - Quaternary) The majority of morphological evidence (and at least one molecular analysis, Gürsoy et al., 2000) supports the monophyly of Vertebrata - lampreys and jawed vertebrates. Characterized by the presence of extensive internal cartilage ("chondrifications")Vertebrate synapomorphies (?):
- Arcualia flanking the spinal cord: Sequentially arranged paired cartilagenous endoskeletal elements (the beginnings of proper ossified vertebrae - right) These, in combination with the notochord, ossify as vertebrae in derived vertebrates. (In contrast, the arcualia of Hyperotreti are below the notochord and only in the tail.)
- Extrinsic eye muscles, allowing eye movements
- Radial muscles in fins, allowing active control of fin position.
- At least two vertical semicircular canals in the otic capsule. But note: Although hagfish have only one pair of semicircular canals (as opposed to two for lampreys), other components of the hagfish otic capsule are paired, and their semicircular canal displays two distinct forms of hair cells, raising the possibility that it represents the evolutionary fusion of two original canals (Heimberg et al. 2010.)
- Lateral line system with neuromasts, sensory capsules enervated by the acoustic nerve.
{kind=link}
{kind=link}
{kind=link}
This is a huge problem in vertebrate systematics that cries out for resolution. The last word is from Miyashita et al., 2019, who present for the first time a morphological analysis containing many fossil taxa that supports the Cyclostome Hypothesis! For the moment, we accept it, but await the fossils that will reveal the true depth of our ignorance.
What we really need is for a Cambrian konzervat-lagerstätte to cough up a fossil basal hagfish or lamprey. Of course, maybe they have and we haven't recognized it. ;-)
- J-Y Chen, J Dzik, G Edgecome, L Ramskold, G-Q Zhou. 1995. A possible early Cambrian chordate. Nature 377, 720-722.
- Jun-Yuan Chen, Di-Ying Huang, Qing-Qing Peng, Hui-Mei Chi, Xiu-Qiang Wang,and Man Feng. 2003. The first tunicate from the Early Cambrian of
South China. Proceedings of the National Academy of Science 100(14) 8314-8318.
- Simon Conway Morris and Jean-Bernard Caron. 2012. Pikaia gracilens Walcott, a stem-group chordate from the Middle Cambrian of British Columbia. Biological Reviews, 87(2): 480-512.
- Frederic Delsuc, Henner Brinkmann, Daniel Chourrout, and Herve Philippe. 2006. Tunicates and not cephalochordates are the closest living relatives of vertebrates. Nature 439, 965-968.
- Ruth Ann Dewel. 2000. Colonial origin for Eumetazoa: major morphological transitions and the origin of bilaterian complexity. Journal of Morphology 243(1): 35-74.
- Diego Garcia-Bellido, Michael Lee, Gregory Edgecombe, James Jago, James Gehling, and John Paterson. 2014. A new vetulicolian from Australia and its bearing on the chordate affinities of an enigmatic Cambrian group. BioMed Central Evolutionary Biology 2014, 14:214 doi:10.1186/s12862-014-0214-z.
- Robert Gess, Michael Coates, and Bruce Rubridge. 2006. A lamprey from the Devonian period of South Africa. Nature 443, 981-984.
- Halil-Cem Gürsoy, Dorota Koper, Bernd-Joachim Benecke. 2000. The Vertebrate 7S K RNA Separates Hagfish (Myxine glutinosa) and Lamprey (Lampetra fluviatilis). Journal of Molecular Evolution, 50(5): 456-464.
- Jian Han, Simon Conway Morris, Qiang Ou, Degan Shu, and Hai Huang. 2017. Meiofaunal deuterostomes from the basal Cambrian of Shaanxi (China). Nature, Published online 30 January 2017.
- Alysha Heimberg, Richard Cowper-Sallari, Marie Semon, Philip Donoghue, Kevin Peterson. 2010. MicroRNAs reveal the interrelationships of hagfish, lampreys, and gnathostomes and the nature of the ancestral vertebrate. Proceedings of the National Academy of Sciences 107(45): 19379Đ19383.
- Nicholas Holland, Grace Panganiban, Erik Henyey, and Linda Holland. 1996. Sequence and developmental expression of AmphiDll, an amphioxus Distalless gene transcribed in the ectoderm, epidermis and nervous system: insights into evolution of craniate forebrain and neural crest. Development 122, 2911-2920.
- Philippe Janvier. 2007. Homologies and Evolutionary Transitions in Early Vertebrate History. In Major Transitions in Vertebrate Evolution. (eds. Hans-Dieter Sues and Jason Anderson), pp. 57 - 121 Indiana University Press, Bloomington.
- Shigehiro Kuraku, Acel Meyer, and Shigeru Kuratani. 2009. Timing of Genome Duplications Relative to the Origin of the Vertebrates: Did Cyclostomes Diverge before or after? Molecular Biology and Evolution. 26(1) 47-59.
- Lacalli, Thurston. 2024. The Cambrian fossil Pikaia, and the origin of chordate somites EvoDevo. Feb., 2024
- Jon Mallatt, J Y Chen. 2003. Fossil sister group of craniates: predicted and found. Journal of Morphology 258(1): 1-31.
- Victoria E. McCoy, Erin E. Saupe, James C. Lamsdell, Lidya G. Tarhan, Sean McMahon, et al. 2016. "The 'Tully monster' is a vertebrate". Nature. 532 (7600): 496Đ499.
- Tetsuo Miyashita. 2018. Ontogeny of Paleozoic stem lampreys as a test of cyclostome-based models of vertebrate ancestry. Society of Vertebrate Paleontology Abstracts, p 183.
- Tetsuto Miyashita, Michael I. Coates, Robert Farrar, Peter Larson, Phillip L. Manning, Roy A. Wogelius, Nicholas P. Edwards, Jennifer AnnŽ, Uwe Bergmann, A. Richard Palmer, and Philip J. Currie. 2019. Hagfish from the Cretaceous Tethys Sea and a reconciliation of the morphologicalĐmolecular conflict in early vertebrate phylogeny. Proceedings of the National Academy of Science published ahead of print January 22, 2019.
- Kinya G. Ota, Satoko Fujimoto, Yasuhiro Oisi, and Shigeru Kuratani. 2011. Identification of vertebra-like elements and their possible differentiation from sclerotomes in the hagfish. Nature Communications 2(373).
- Imran Rahman, Samuel Zamora, Peter Falkingham, and Jeremy Phillips. 2015. Cambrian cinctan echinoderms shed light on feeding in the ancestral deuterostome. Proceedings of the Royal Society. B 282: 20151964. http://dx.doi.org/10.1098/rspb.2015.1964.
- Lauren Sallan, Sam Giles, Robert S. Sansom, John T. Clarke, Zerina Johanson, Ivan J. Sansom, Philippe Janvier. 2017. The ÔTully MonsterŐ is not a vertebrate: characters, convergence and taphonomy in Palaeozoic problematic animals. Palaeontology 60(2): 149-157.
- Michael Schubert, Hector Escriva, José Xavier-Neto, Vincent Laudet. 2006. Amphioxus and tunicates as evolutionary model systems. Trends in Ecology and Evolution, 21 - 5, 269 - 277.
- Degan Shu. 2003. A paleontological perspective of vertebrate origin. Chinese Science Bulletin 48(8): 725-735.
- Degan Shu, X Zhang, L Chen. 1996. Reinterpretation of Yunnanozoon as the earliest known hemichordate. Nature 380, 428-430.
- D Shu, S Conway Morris, J Han, L Chen, X Zhang, Z Zhang, H Liu, Y Li, and J Liu. 2001. Primitive deuterostomes from the Chengjiang Lagerstätte (Lower Cambrian, China). Nature 414, 419-424.
- D-G Shu, L Chen, J Han, and X Zhang. 2001. An Early Cambrian tunicate from China. Nature 411, 472-473.
- Billie Swalla, Chris Cameron, Laura Corley,a nd James Garey. 2000. Urochordates Are Monophyletic Within the Deuterostomes. Systematic Biology 49(1): 52-64.
- Qingyi Tian, Fangchen Zhao, Han Zeng, Maoyan Zhu, and Baoyu Jiang. 2022.
Ultrastructure reveals ancestral vertebrate pharyngeal skeleton in yunnanozoans. Science, 377(6602), 218 - 222.- Jacob Vinther, M. Paul Smith, David A. T Harper. 2011. Vetulicolians from the Lower Cambrian Sirius Passet Lagerstätte, North Greenland, and the polarity of morphological characters in basal deuterostomes. Palaeontology, 54 - 3, 711 - 719.
- D. Yalden, 2008. Feeding mechanisms as evidence for cyclostome monophyly. Zoological Journal of the Linnean Society. 84(3): 291-300.
- Simon Conway Morris and Jean-Bernard Caron. 2012. Pikaia gracilens Walcott, a stem-group chordate from the Middle Cambrian of British Columbia. Biological Reviews, 87(2): 480-512.