

Deuterostomia III: Chordata
Chordata:
The sister taxon of Ambulacraria. Contains:- Urochordata: Tunicates (aka "sea-squirts")
- Cephalochordata: Living Branchiostoma (aka amphioxus, aka lancelets. Deal with it.)
- Craniata: Creatures with vertebrate-style heads - hagfish, proper vertebrates, and a few interesting fossil taxa.

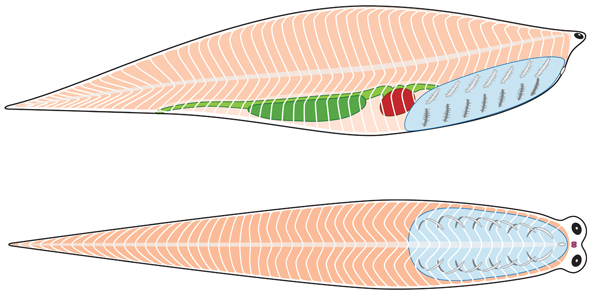
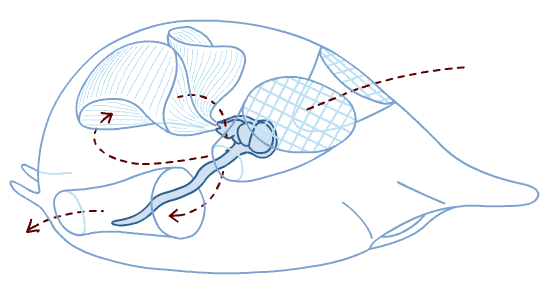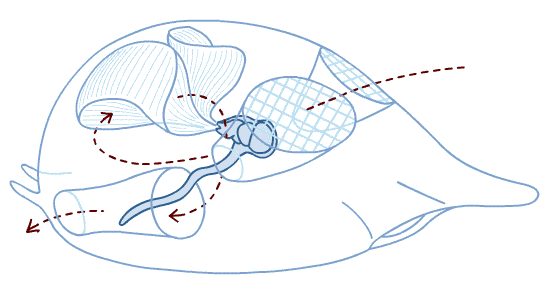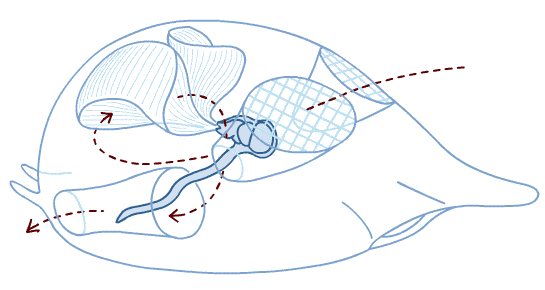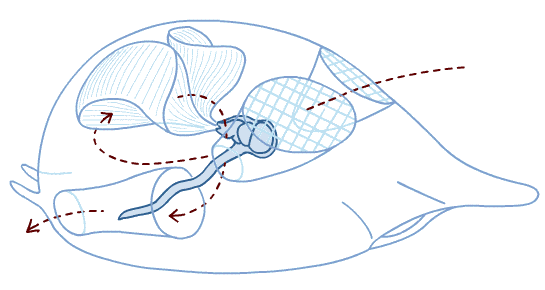
Schematic of ancestral vertebrate (showing chordate characters plus some)
from Radinsky 1987. The Evolution of vertebrate Design.
- Body stiffened by a notochord - a rod made of cartilage-like tissue and fluid wrapped in a tight membrane - at some point in life
- Dorsal nerve cord hollow for entire length.
- Mucus-secreting endostyle at ventral midline of pharynx.
- Post-anal tail.
- Body musculature divided into myomeres
Pikaia - Almost a chordate:

The Burgess Shale fossil Pikaia graciliens, known from hundreds of specimens, is often cited in popular literature as the "earliest chordate." In fact:
- It is not the earliest. That would be either Myllokunmyngia, the similar Haikouicthys, or Cathaymyrus from Chengjiang.
- Based on a recent redescription by Conway-Morris and Caron, 2012, it is not quite a chordate as it lacks a post-anal tail or urostyle, and its notochord (if that's what it is) is rudimentary. Link to interpretation.
- Chevron-shaped myomeres
- Mid-line dorsal and ventral fins
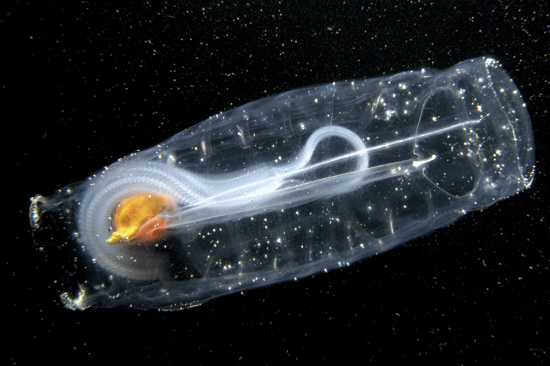
Cephalochordata: (Cam. - Rec.) "Headless fish."

Represented by the living Branchiostoma.
Confusing nomenclature: Earlier names have priority in scientific nomenclature. Unfortunately, the publication in which this animal was originally named (Branchiostoma) was largely ignored, and it was named a second time (Amphioxus). Additionally, there had been a common name (Lancelet). When the dust settled, Branchiostoma was acknowledged the proper scientific name, and amphioxus and lancelet are both considered common names. Live with it.
Anatomy: In many ways, Branchiostoma is like a modified version of Pikaia. Similarities include:
- Food is captured by filtration of water propelled through a pharynx by ciliary action.
- Muscles enabling the animal to swim by lateral undulations are divided in to chevron-shaped myomeres.
- Dorsal nerve cord.
- Mid-line dorsal and ventral fins are present forming a tail.

Where Branchiostoma differs significantly:
- Body supported by a robust notochord that extends from the tip of the snout to the end of the tail. This supports...
- The dorsal nerve cord, which is hollow for its entire length.
- A distinct oral chamber is separated from the pharynx by a velum
- the pharynx is:
- Large
- Pharyngeal openings are tall slits separated by pharyngeal bars.
- Water filtered through pharyngeal slits by ciliary action enters an atrium - an outer chamber that envelops the pharynx, before it is expelled.
- A gland called the endostyle occupies the ventral mid-line of the pharynx. It secretes mucus that is carried along the pharyngeal bars by cilia. This mucus, along with anything digestible that sticks to it, is transported to the esophagus and gut tube.
- The post-anal tail tail extends well past the anus.
Branchiostoma ecology: Branchiostoma opens a window onto the ecology of the "swimming pharynges" at the base of Deuterostomia. It does not spend much time actually swimming, but, instead, buries itself in soft sediment, with its front end projecting into the water suspension-feeding. When disturbed, it swims in a fish-like manner, but soon settles down, digs in, and resumes suspension-feeding.

Anatomical details:
- The dorsal nerve cord is hollow and there is a slight anterior enlargement. (Indeed, genes expressed in vertebrate brains are also expressed in this region, providing some basis for the establishment of homology.
- A diverticulum of the gut occupies the place of the liver in craniates.
Synapomorphy of Cephalochordata:
- Notochord extends to the front end of the body.
Cephalochordate fossil record: Sparse, but present as early as the Chengjiang lagerstätte in the form of Cathaymyrus diadexus.
Interpreting cephalochordates: For decades, cephalochordates have been presumed to be closely related to Chordata. They are so close morphologically. The Wizard need only give them hearts and brains. The next group, in contrast, seems more distant.
Urochordata: (Sparse record Cam. - Rec.)

Also known as "tunicates" or "sea squirts." As adults, the body is dominated by a huge pharynx used for suspension-feeding. Adults of readily accessible species are typically attached to some hard surface, however many are free-swimming. They may be solitary or colonial.
Anatomy:
- Their bodies consist of a large basket-like pharynx perforated by numerous slits. On one side, the endostyle secretes mucus which is transported across the inner surface of the pharynx by cilia, capturing food particles as it goes.
- Entire body enclosed by a "tunic" made primarily of tunicin, a unique complex sugar. Water enters pharynx through a "mouth" and, after filtration, enters an atrium from which it is expelled through a separate opening, the atriopore, as in Branchiostoma.
- Unlike Branchiostoma, the anus also opens into the atrium,
- a notochord
- a hollow dorsal nerve cord
- muscles arranged into myomeres
Diversity: Traditionally divided into three groups.
-
Ascidiacea: Spend their adult lives attached to an accessible-to-biologists shallow marine substrate. May be solitary (right) or colonial (see above). Their accessibility makes them the standard urochordate of textbooks and field guides, coloring our perceptions of the entire group.
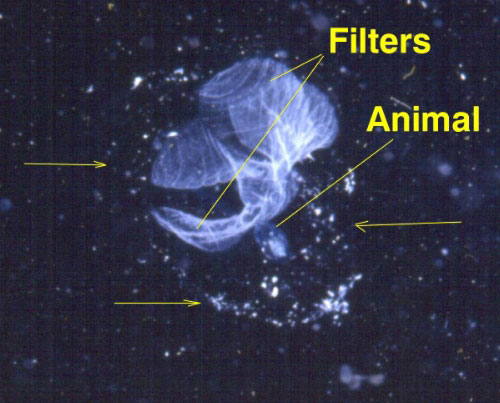
- Thaliacea: Salps. Metamorphose into cylindrical living pharynxes but do not attach to the substrate. Incurrent and excurrent openings at opposite ends of the body. The tunic is invested with muscle, alowing them to swim by "jet-propulsion." Can be solitary or colonial.

Larvacea: Metamorphose from non-feeding tadpoles to feeding adults, but this change is less extreme than in other urochordates as they retain their tails, notochords, and nerve cords. The freaky stuff - Adults secrete a gelatinous structure, the house (that's the technical term) which they inhabit and push around as they swim. The house gathers and concentrates food in suspension. When it becomes fouled, the animal abandons it and secretes a new one. Invariably solitary.


Urochordate evolution: We have two sources of information:
- A sparse fossil record from Chengjiang: Shanlouclava shankouense (right) from Chen et al., 2003 and Cheungkongella from Shu et al. 2001 b have been proposed as fossil urochordates. Each claim has its proponents and critics. As reconstructed, Shankouclava looks like an ascidiacean that never completely resorbed its tail.
- But Garcia-Bellido et al., 2014 regard vetulicolians as stem urochordates.


- Larvacea is the basal branch of urochordata
- Ascidiacea is paraphyletic with respect to Thaliacea
- The colonial lifestyle evolved several times independently.

When your instructors completed their PhDs, the consensus of deuterostome phylogenies held that:
- Hemichordata was the sister taxon to Chordata. Synapomorphy: Pharyngeal openings.
- Cephalochordata was the sister taxon of Craniata. Synapomorphies:
- The Burgess shale fossil Pikaia was a cephalochordate because it resembled Branchiostoma and had no distinct head.
- Urochordates we considered to represent an ancestral form, from which cephalochordates and craniates might have evolved by pedomorphosis.
Discoveries that have changed that view:
- Vetulicolians and Yunnanozoans provide a picture of the ancestral deuterostome as a "swimming pharynx." Thus:
- The swimming morphology and segmentation of craniates, cephalochordates, larval urochordates, and Pikaia are plesiomorphies.
- Same for the presence of pharyngeal openings in hemichrodates and chordates.
- If Larvacea represents the basal condition for Urochordata (Swalla et al. 2000) then urochordates also arose from "swimming phayrnges." (Indeed, Garcia-Bellido et al. 2014 propose that vetulicolians are actually stem urochordates. We await confirmation.)
- Strong molecular evidence for Olfactores and Ambulacraria that doesn't imply improbable reversals and convergences.
- Re-description and "demotion" of Pikaia.
"Craniates" - Chordates with heads:
The remaining chordates are strongly distinct creatures. Synapomorphies include:- Head including:
- Anterior expansion of nerve cord into brain.
- Special sense organs:
- Olfactory capsules for olfaction
- Eyes for vision
- Otic capsules for hearing
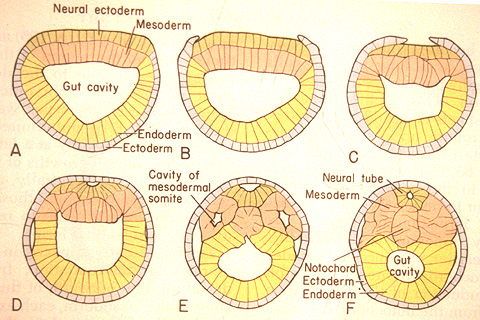
Neurulation in Branchiostoma from A. S. Romer. 1977. The Vertebrate Body.
Chordate Development:
Neurulation: The indeterminate nature of deuterostome development, in which the fates of specific cells are influenced by inductive relationships with other cells, is illustrated by the next big step chordate development - the formation of the neural tube that gives rise to the central nervous system. In this process, the activity of mesoderm cells triggers a cascade of events.- Cells on the mid line of the mesoderm form a cylinder that becomes the notochord.
- Mesoderm to either side folds outward and expands down the side of the gastrula's inner surface.
- An inductive relationship causes the ectoderm directly above the notochord to sink downward, forming a trough, and ectoderm to either side to rise up forming crests to either side. The trough collapses into a hollow tube lying above the precursor to the notochord. This is the neural tube - the precursor to the brain and spinal cord.
The special sense organs arise from the interaction of the neural tube with placodes of the outer ectoderm.
Additional derived features:
- Neural crest tissue forms significant portions of the head, gill-arch skeleton, peripheral nervous system, and pigmentation.
- Water pumped through pharynx by muscles instead of my ciliary action.
- The bars separating the slits of the pharynx support special gas-exchange organs - gills.
- Cartilaginous fin-rays support the fins.
- Arcualia - cartilaginous precursors to vertebrae.
- W - shaped myomeres - an elaboration of the simple chevrons of Chordata.

- Orders of magnitude bigger than any non-craniate chordate
- Much more active than non-craniate chordates.
Olfactores - Urochordate - craniate synapomorphies:
- although according to Holland et al., 1996, neural crest homologs (marked by gene expression) may be present in Branchiostoma, morphologically identifiable neural crest is absent, as are ectodermal placodes.
- In urochordates, migratory neural crest cells are present, but only control the distribution of pigment in the skin. Although there are no special sense organs, the atrium arises through the invagination of an ectodermal placode.
Schematic of ancestral vertebrate (showing chordate characters plus some)
from Radinsky 1987. The Evolution of vertebrate Design.
- Mouth, with more specialized mouth parts
- Arches of pharynx now support gills, used for both feeding and respiration
- Branchial apparatus supported by cartiligenous brachial arches (gill bars) external to gills
- Pharyngeal muscles pump water through pharynx
- Caudal fin stregthened by cartiligenous radials
- Larval endostyle transforms into adult thyroid gland
- In the gut:
- Liver and pancreas present (formed through endoderm-mesoderm induction)
- Digestive system invested with smooth muscular lining (rather than cilia)
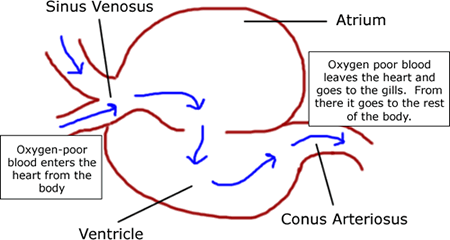
- Circulatory System
- Two chambered heart
- Hemoglobin for oxygen transport
- Erythrocytes (red blood cells) to contain hemoglobin
- Increased size (an order of magnitude)
- Cartilaginous endoskeleton: including fin rays and brachial arches
In terms of metabolic rate and aerobic capacity, only cephalopod mollusks and some arthropods are comparable to craniates. Major groups:
- Hyperotreti
- Hyperoartia
- Gnathostomata
Craniate diversity:
First, consider the living groups:

Hyperotreti:
(Hagfish - Carboniferous - Quaternary). Synapomorphies of Hyperotreti:- Comb-shaped biting mouthparts
- Four pairs of tentacles surround the mouth.
- Impressive mucus glands
Other characteristics:
- The skeleton consists of the notochord and specialized cartilages of mouth. The latter take the form of a mid-line rod-and-pulley upon which the tooth-plates are protracted and retracted.
- Otic (inner ear) capsules have only one semicircular canal (as opposed to three in jawed vertebrates).
- An adult hagfish is much too big to achieve gas exchange by simple diffusion. Its gill slits are, therefore, lined with thin pleats of heavily vascularized tissue - proper gills - which serve as breathing organs.
- Five to fifteen pairs of gill openings.
But hagfish lack many features we see in other craniates:
- Any suggestion of arcualia outside of the tail. (In hagfish, the only arcualia form in the position of the hemal arches of more derived vertebrates.)
- A brain-case as part of their skeleton. Theirs is largely membranous.
- Extrinsic eye muscles: Hagfish eyes are simple and cannot be rotated in the head.

Hyperoartia:
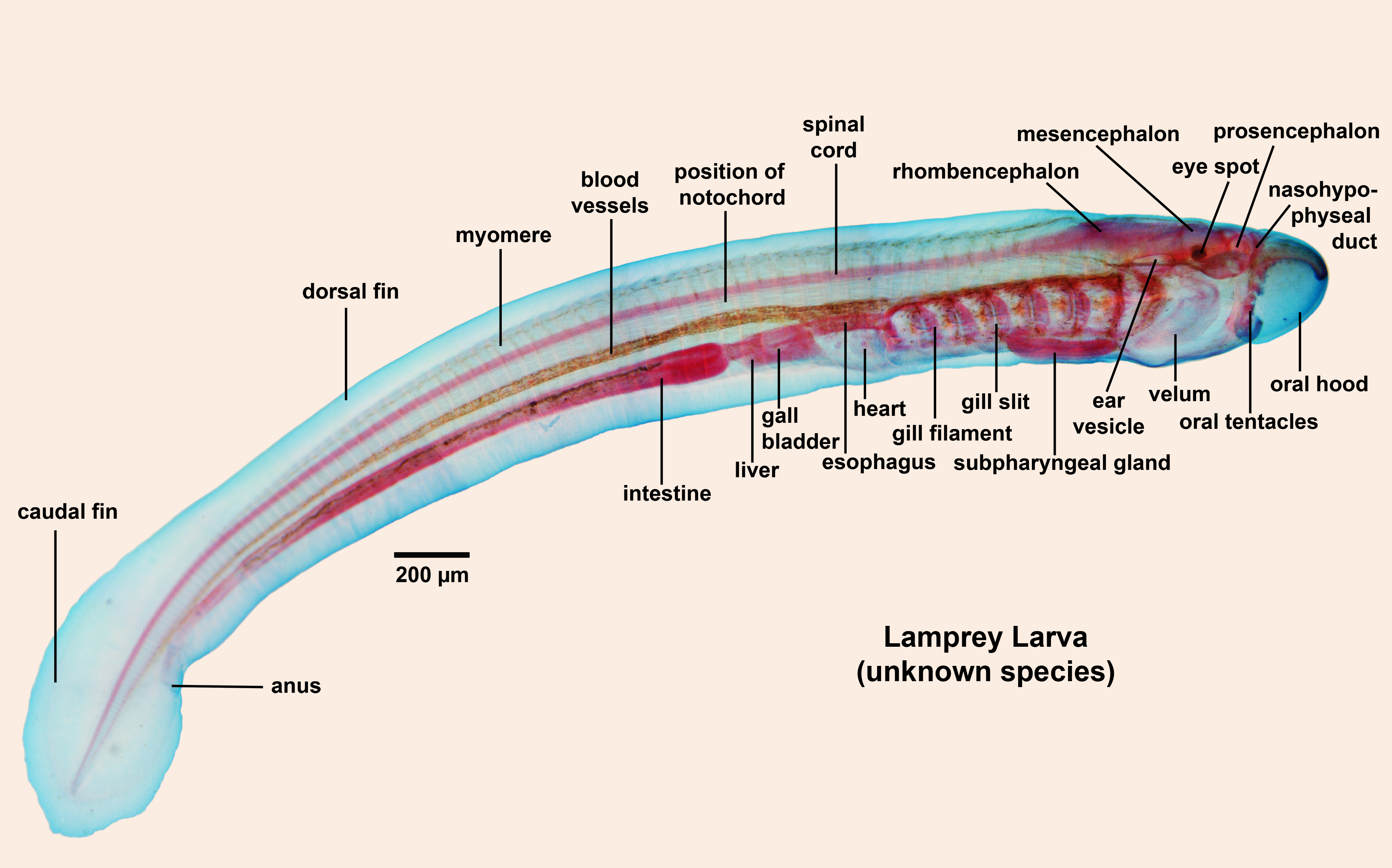
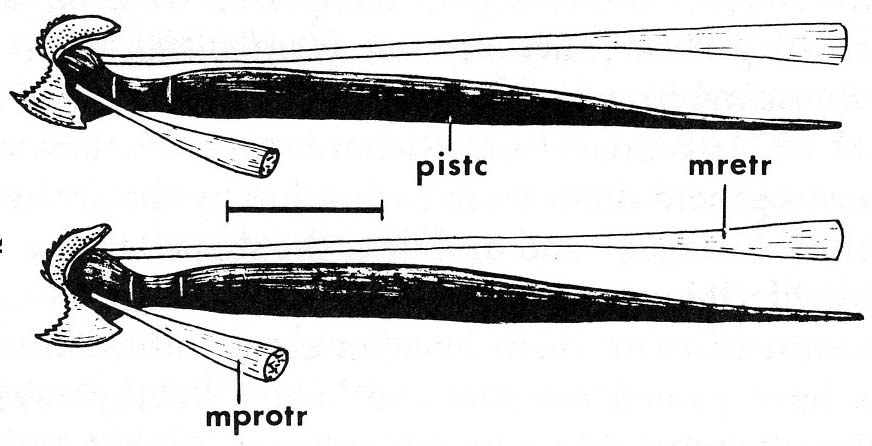
(AKA Petromyzontida, aka lampreys) - (Devonian - Recent): Characterized by:- Undergoes metamorphosis from suspension-feeding ammocoetes larva that looks like a brainy version of Branchiostoma to a parasitic adult.
- In adult, annular cartilage supports a large sucker surrounding the mouth armed with keratin "teeth".
- Piston cartilage supporting a protrusible "tongue" armed with more keratinous denticles. Although distinct, this is also a rod-and-pulley arrangement similar to that of hagfish.
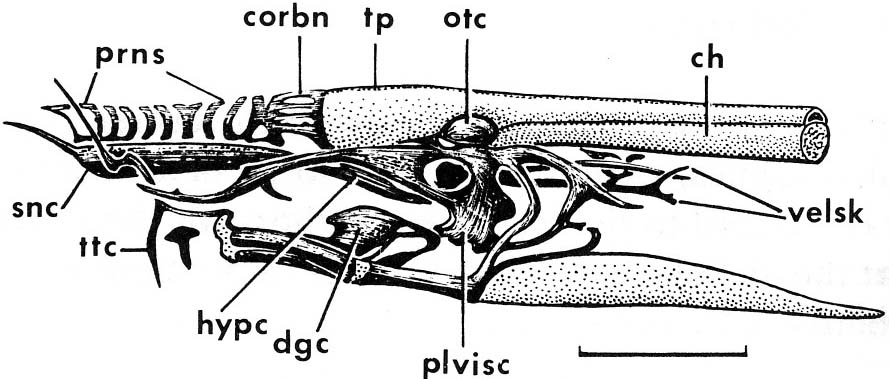
- Branchial skeleton: the lamprey gill pouches are supported by a branchial basket whose cartilagenous elements are external to the gills.
- Seven pairs of gill openings.
- Tail bends downward slightly (barely hypocercal)
- Living taxa have two distinct dorsal fins, although fossils don't.
- Two vertical semicircular canals in the otic capsule. (In this and in other respects considered below, lampreys appear to have much in common with the final craniate group.)

Priscomayon riniensis (Late Devonian) and Lampreta fluviatilis (Quaternary)

Gnathostomata:
(Silurian - Quaternary) The last common ancestor of jawed vertebrates and all of its descendants. Living gnathostomes are distinct from living either Hyperotreti or Hyperoartia in many respects. Conspicuously, they have jaws, but also many other features to be discussed later.
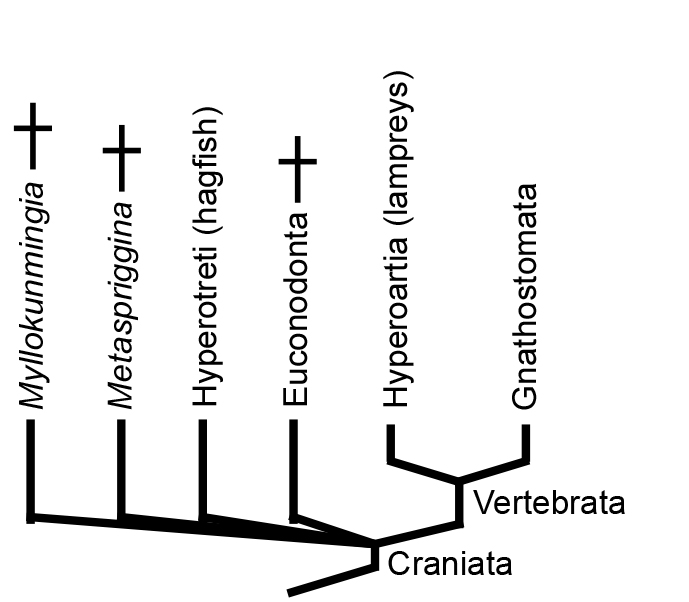
"Craniate" Relationships
Illuminating the relationship between Hyperotreti, Hyperoartia, and Gnathostomata is difficult and contentious. Two major hypotheses predominate that take their names from the positions they imply for lampreys:
- The Cyclostomata hypothesis, in which Hyperotreti and Hyperoartia form a clade. Together, Cyclostomata and Gnathostomata make up Vertebrata (of which "Craniata" would be a junior synonym).
- The Vertebrata hypothesis, in which Hyperoartia group with Gnathostomata - Vertebrata. Together, Vertebrata and Hypertreti make up a monophyletic Craniata.
This is a huge problem in vertebrate systematics that cries out for resolution. Janvier's quote is apt, but opinion is swinging toward the Cyclostomata hyopthesis. What we really need is for a Cambrian konzervat-lagerstätte to cough up a fossil basal hagfish or lamprey. Of course, maybe they have and we haven't recognized it. ;-)
Hints of unseen diversity - conodonts

Conodont elements
- Highly diverse and rapidly evolving, thus excellent index fossils.
- Originally proposed to be the teeth of some unknown fish, but paleontologists soon determined they were were clueless about:
- What kind of animal they were from
- What part of the animal they represented.
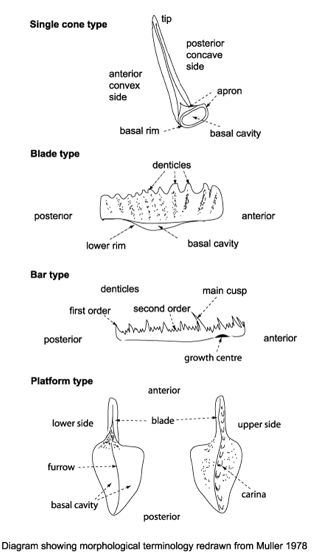
Conodont types
At this point, speculation raged about:
- Which major taxon the elements belonged to
- How were they used? Originally proposed to be dental elements of fish-like chordates by Pander (1856), but soon, more erudite-sounding hypotheses held sway.
- To some they seemed likely to be used in prey capture, as in similar sturctures in chaetognaths (arrow-worms)
- To others, they seemed like mineralizations of the suspension feeding apparatus of cephalochordates.
- Or perhaps they were for internal support and not for feeding at all.

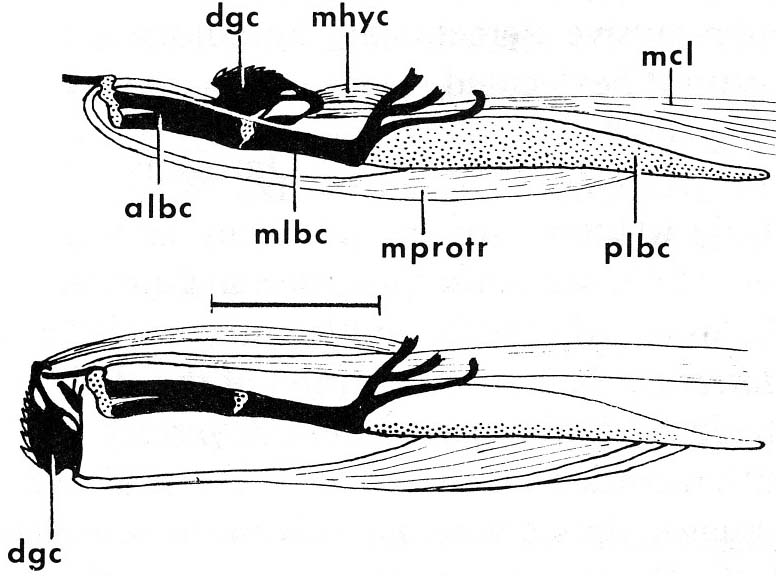
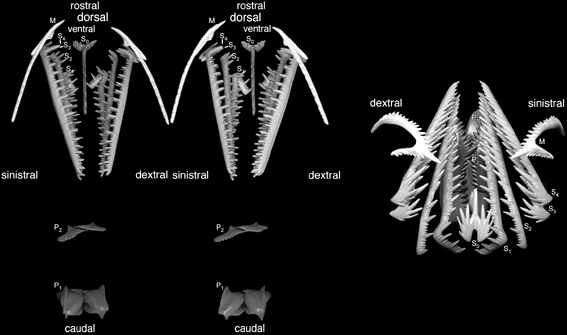
Conodont apparatus
Since these seemed to appear in a stratigraphic (chronological) sequence, speculation was that these growth forms represented evolutionary stages. Not quite. In 1988, single cone elements were eliminated from the roster of euconodonts by the discovery that they were the fossils of early chaetognaths. In living chaetognaths, similar elements are made of calcium carbonate and are used in prey capture.
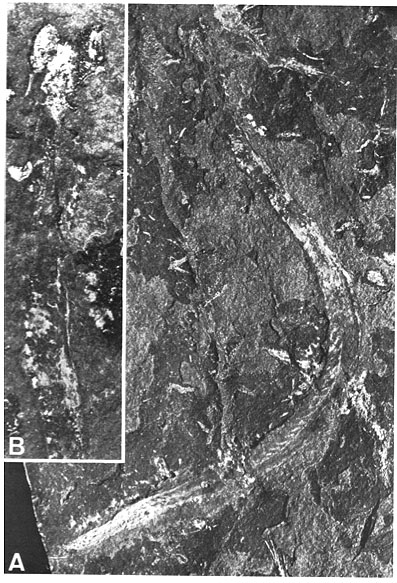
Clydagnathus
- Chordate-like V-shaped segmented muscle blocks
- Midline fins supported by fin rays
- The conodont apparatus in an anterior position, suitable for use in feeding.
- Notochord
- A head a brain and two capsules for special senses, thought to be very large eyes and smaller otic capsules.

Euconodonts
But where does Euconodonta go on the chordate cladogram? The presence of phosphatic hard parts arguably places it, closer to Gnathostomata than to hagfish or lampreys, but there are concerns:
- The conodont elements, as elements, are not homologous to any other phosphatic vertebrate feature.
- Jawless vertebrates might have secondarily lost hard tissues as a reversal.
- Some morphological interpretations of euconodonts, especially the huge eyes, seem to cry out for revision.
- The assumption of euconodont monophyly hasn't really been rigorously tested.
The earliest vertebrates:
Chengjiang and the Burgess Shale have provided other records of basal vertebrates lacking conodont elements.Myllokunmingia and Haikouichthys: Chengjiang gives us the best picture picture of what the ancestral vertebrate might have looked like with Haikouichthys ercaicunensis (right - possibly junior synonym of Myllokunmingia fengjiaoa), which seem to preserve:

- W-shaped myomeres
- placode-derived special sense organs
- a branchial skeleton resembling that of lampreys
- Dorsal fin with possible fin rays
- possible arcualia the anterior notochord
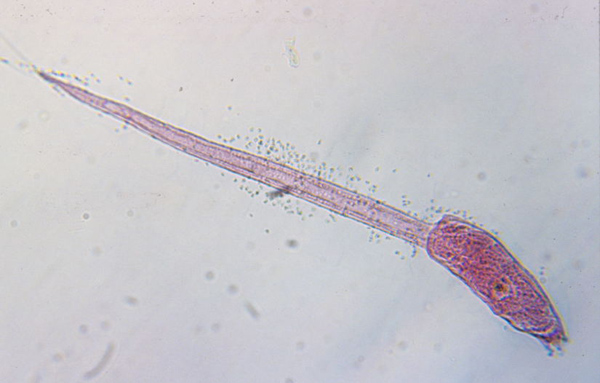
Metaspriggina: And from the Burgess shale, Conway-Morris and Caron, 2014 describe Metaspriggina walcotti (right). In this case, the presence of eucondont-like eyes and W-shaped myomeres is unambiguous.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
- Derek Briggs, Euan Clarkson, and Richard Aldridge. 1983, The Conodont Animal. Lethaea, 16(1), 1-14.
- Jun-Yuan Chen, Di-Ying Huang, Qing-Qing Peng, Hui-Mei Chi, Xiu-Qiang Wang,and Man Feng. 2003. The first tunicate from the Early Cambrian of South China. Proceedings of the National Academy of Science 100(14) 8314-8318.
- Simon Conway Morris and Jean-Bernard Caron. 2012. Pikaia gracilens Walcott, a stem-group chordate from the Middle Cambrian of British Columbia. Biological Reviews, 87(2): 480-512.
- Simon Conway Morris and Jean-Bernard Caron. 2014. A primitive fish from the Cambrian of North America. 512, 419-422.
- Frederic Delsuc, Henner Brinkmann, Daniel Chourrout, and Herve Philippe. 2006. Tunicates and not cephalochordates are the closest living relatives of vertebrates. Nature 439, 965-968.
- Nicolas Goudemand, Machael Orchard, Severine Urdy, Hugo Bucher, and Paul Tafforeau. 2011, Synchrotron-aided reconstruction of the conodont feeding apparatus and implications for the mouth of the first vertebrates. Proceedings of the National Academy of Sciences, 108(21), 8720-8724.
- Alysha Heimberg, Richard Cowper-Sallari, Marie Semon, Philip Donoghue, Kevin Peterson. 2010. MicroRNAs reveal the interrelationships of hagfish, lampreys, and gnathostomes and the nature of the ancestral vertebrate. Proceedings of the National Academy of Sciences 107(45): 19379-19383.
- Nicholas Holland, Grace Panganiban, Erik Henyey, and Linda Holland. 1996. Sequence and developmental expression of AmphiDll, an amphioxus Distalless gene transcribed in the ectoderm, epidermis and nervous system: insights into evolution of craniate forebrain and neural crest. Development 122, 2911-2920.
- D-G Shu, L Chen, J Han, and X Zhang. 2001. An Early Cambrian tunicate from China. Nature 411, 472-473.
- Billie Swalla, Chris Cameron, Laura Corley,a nd James Garey. 2000. Urochordates Are Monophyletic Within the Deuterostomes. Systematic Biology 49(1): 52-64.
- D. Yalden, 2008. Feeding mechanisms as evidence for cyclostome monophyly. Zoological Journal of the Linnean Society. 84(3): 291-300.
To Next Lecture.
To Previous Lecture.
To Syllabus.