GEOL 331 Invertebrate Paleontology
Fall Semester 2008
Embryophyta - land plants
We care about land plant, or embryophyte, evolution for many reasons:
- Plants are the primary producers for most ecosystems.
- Plants form the framework of the physical environment in which land animals live. In fact, in the history of life on land, plants have functioned as pioneers, evolving to live in new environments, and making those environments hospitable for animals.
- Plants influence sedimentation
- The carbonization and exhumation of plants influences the carbon cycle.
- Plants have a copious fossil record. Indeed, in the form of coal, they are economically important.
- Land plant fossils, both macro and microscopic, are ubiquitous and precise paleoenvironmental indicators - the forams of the land.
Plant origins
We must revisit an issue that we took up at the beginning of our survey of the diversity of fossil forming organisms: Endosymbiosis.
- As far as we know, oxidative photosynthesis evolved only once, in cyanobacteria. (Although anoxic photoautotrophs use different electron donors)
- Chlorophyll: The primary photosnthetic pigment, although other pigments can be involved.
- Plastids: A family of organelles of photosynthesizers, including chloroplasts, responsible for photosynthesis. These arose from the endosymbiosis of cyanobacteria.
- Secondary endosymbiosis: The incorporation of photosynthesizing eukaryotes by endosymbiosis has enabled several distinct eukaryote lineages to become photosynthesizers.
- Plants sensu lato are descendants of the first organisms with plastids - i.e. the products of the primary endosymbiosis of cyanobacteria.
|
Plant phylogeny:
- Glaucophyta: (No record) Most primitive, unicellular
- Rhodophyta: (Red algae - Neoproterozoic - Rec.) Discussed in previous lecture. Unicellular and multicellular. Exclusively marine
Many are coralline - i.e secrete calcareous skeletons.
- Chlorophyta: Unicellular and multicellular, marine and freshwater.
- Mesostigma: Fresh water unicellular.
- Charophyta Multicellular aquatic. Paraphyletic. Note that these plants do not alternate generations, and produce sperm and ova, although the sperm may not have flagella.
- Charales Multicellular aquatic.
- Embryophyta land plants.
|

|
Adaptations to life on land:
The closest sister taxa to land plants are the paraphyletic grade group of multicellular green algae known as charophytes Charophytes:
- exchange gasses with the environment by simple diffusion,
- require no structural tissue to support their bodies,
- release their sperm (and in some, ova) directly into the water.
In order for plants to colonize the land, they had to overcome the challenges of retaining water, exchanging gasses, supporting their bodies, and reproducing out of water. Five adaptations facilitated their response:
- A waterproof cuticle covering most of the external surface prevents desiccation.
- A vascular system of fluid conducting cells to move liquids through the plant.
- Stomata, controllable microscopic openings allowing gas exchange through the impermeable cuticle. (These open and close in response to illumination, humidity, and [CO2].
- Gametes no good for dispersal on land, hence dispersal through spores with durable walls to resist desiccation.
- Were do we get spores? Alternation of generations in which the parent gametophyte shelters and supports a growing sporophyte.
Alternation of Generations:
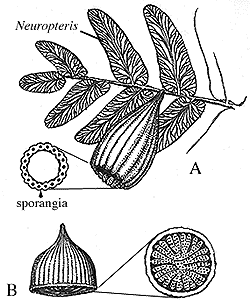
- Adult diploid sporophytes produce haploid spores. These usually develop in capsules called sporangia which break open, allowing the spores to disperse.
- Spores grow into haploid gametophytes. These produce male and female gametes. In more primitive embryophytes, regions of the gametophyte known as antheridia and archegonia produce sperm and ova respectively. The sperm swim to the through a film of water on the plant's surface ovum to form a diploid zygote.
- The need for the sperm to reach the ova under their own power imposed a powerful constraint on the earliest embryophytes: Their gametophyte generation had to grow in moist environments.
- The zygote growing from the archegonium, develops into a new diploid sporophyte. Sporophytes generally differ from gametophytes in possessing tall stems. Why? Presumably to elevate the sporangia for better spore dispersal.
Apparently, in the ancestral embryophyte, the gametophyte generation was emphasized and the sporophyte generation was relatively ephemeral. Because of the gametes dependency on water, such plants could only thrive in moist environments. Plants of this sort, including liverworts, mosses, and hornworts, were traditionally classed in the paraphyletic "Bryophyta."
Tracheophyta:
The tracheophytes, or "vascular plants" include the majority of land plants. As the name implies, they are characterized by vascular tissue reinforced by lignin, a durable substance contributing to vascular tissue and call walls. The presence of this substance facilitated the growth of taller, stronger stems. This was a vital prerequisite to their second major adaptation: The emphasizing of the sporophyte generation over the gametophyte generation.
Modern ferns give a good idea of the ancestral tracheophyte reproductive system. Their gametophytes are generally only a few millimeters across, whereas their sporophytes can be quite large. By means of this innovation, ferns and other tracheophytes can break their dependence on moist environments for most of their life cycles. Although their gametophytes still require moisture, they are small enough that they can develop from spores quickly when favorable conditions occur.
The bodies of vascular plant sporophytes are generally differentiated into:
- rhizomes which run along the ground, connecting individual plants,
- roots, which obtain minerals from the soil, and
- stems, which produce glucose through photosynthesis above the ground.
- In all but the most primitive, leaves increase the amount of sunlight that can be captured.
Vascular tissue allows nutrients from the soil and the glucose derived from photosynthesis to be delivered to the entire plant.
"Seedless vascular plants": The first major radiation of vascular plants occurred in the Silurian and Early Devonian, giving rise to groups that dominated land floras for most of the Paleozoic This radiation consisted primarily of plants that reproduced in the manner described above:
- Rhyniophytes:" Sil. - Dev. The paraphyletic group containing the earliest and most primitive kown tracheophytes is named after a well-known genus, Rhynia. These plants consisted of rhizomes and leafless stems with small simple roots, were generally less than 10 cm. tall, and bore sporangia at the tips of their stems. The leafless modern Psilotum superficially resembles them, and was once thought to be a surviving member of this early radiation, but it is now thought to be a highly derived fern.
- Lycophyta: (Lycopods) Dev. - rec.
Members of this group possess narrow pointed leaves that are vascularized by a single strand of vascular tissue. The leaves cover the stem. When they fall away, they leave the stem covered by characteristic diamond shaped scars. Modern representatives, the "club mosses," are a sad remnant of a once mighty radiation. During the Mississippian and Pennsylvanian, their tree-sized relatives, including Lepididendron and Sigillaria were the largest forest plants. The sporangia of lycophytes are born on the upper surface of specialized leaves which grow in groups, forming cones or strobili at the ends of stems.
Earliest representatives of lycophyte lineage may be Zosterophyllopsida, (Zosterophyls) resembling rhyniophytes but with alternating sporangia on stem.
|

|
- Monilophyta (ferns and horsetails): Late Dev. - rec.
Ferns were the first plants with large vascularized leaves. Most, but not all ferns have very short trunks that support large pinnate leaves. Many ferns have independently evolved the "tree-fern" habitus, growing to the size of small trees. Post-paleozoic tree ferns seem to be monophyletic, but there were paleozoic ferns that separately evolved the tree-fern form. Ferns are plesiomorphic in bearing their sporangia on the bottoms of unmodified leaves.
|

|
|
One monilophyte grope has only recently been found to group within "ferns" - the Sphenopida or "horse-tails. (Dev. - Rec). Like lycophytes, these plants protect their sporangia in strobili that develop at the end of stems. They are distinguished by the unique morphology of their stems. These consist of cylindrical sections separated by nodes. Whorles of slender leaves radiate from the nodes. During the late Paleozoic and Triassic, some sphenopsids such as Calamites attained tree size, but they were never the primary flora of their environments. Modern sphenopsids are known for rendering themselves inpalatable by incorporating silica phytoliths into their tissues.
|

|
Spermatophyta - Seed Plants:
Seeds: The second great radiation of land plants occurred during the Late Paleozoic, and was associated with the evolution of the seed. This occurred in stages.
- Ancestrally, gametophytes produced both sperm and ova.
- Some fossil as well as some recent non-seed bearing plants, display a dimorphism of between megaspores and microspores, which specialize in producing either ova or sperm, respectively and which resemble them in size dimorphism. This trend seems to develop primarily in plants with strobili which can, in principle, harbor the gametophyte as well as the sporangia.
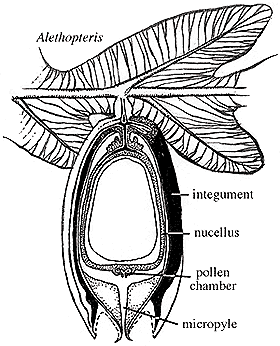
- When we pick up this trend again, it is in plants that retain the female gametophyte inside special reproductive structures. The gametophyte is known as an ovule, and contains the ovum, nutritive tissue, a seed coat, and structures for conducting sperm to the ovum. Once the ovum is fertilized, the ovule becomes a seed.
- How does the sperm get to the ovum? When a microspore lands on the sperm-capturing portion of the ovule, it develops into a specialized male gametophyte called a pollen-tube which grows into the ovule. Sperm is conducted down the pollen-tube to the ovum. In such a set-up, we call the microspore a pollen grain.
Wood: A second major evolutionary novelty was the ability of seed plants to lay down secondary tissue. Ancestrally, land plants could create new tissue only in the apical meristem tissue at the tips of their stems. In seed plants, however, we see the appearance of vascular cambium, a sheath of tissue that generated new vascular tissue around the circumference of the stem. New tissue laid down by vascular cambuim appears as growth rings in cross sectioned stems. The ability to thicken existing stems allowed seed plants to attain greater size than their arborescent lycopod precursors.
"Progymnosperms" Paraphyletic grade group of plants bearing vascular cambium but lacking ovule-like reproductive structures:
- Archaeopteridales: (Devonian - Mississippian) Archaeopteris, is typical. Major component of the Late Devonian forests, including the first large (redwood-sized) trees. Ovules and pollen organs were absent, but displays distinction between mega and micro spores.
|

|
"Seed ferns" Paraphyletic grade group including numerous Late Paleozoic land plants characterized by distinct ovules and pollen organs born on separate leaves:
Spermatophyta: The crown-group of seed plants. Contains five well-diagnosed monophyletic groups:
- Conifers: P. - rec. carry their seeds on the scales of cones. Most conifers are tree-sized and produce both male (pollen bearing) and female (seed bearing) cones. In many cases, their leaves are reduced to needles, allowing them to withstand cold and dry conditions. This ability allows many to retain their leaves year-round. Earliest known representative were Late Paleozoic Cordaitales, trees with whorls of strap-like leaves. Living examples:
|

|
- Ginkgoales (Ginkgos) P. - rec. Common in the Mesozoic and Paleogene, they are now reduced to a single species, Ginkgo biloba. Like cycads, ginkgos are dioecious - have separate male and female individuals. The ovule is enclosed in a malodorous fleshy capsule.
Represented by a single species, Ginkgo biloba, but speciose during the Mesozoic.
|

|
- Cycadales: (Cycads) Penn. - rec. Shrub-sized superficially fern or palm-like plants with short trunks and very tough pinnate leaves. They are very slow-growing. Like conifers, they produce male and female strobili. In this case, however, each individual plant is fully male or female, producing cones of only one sex. Cycads were very common understory plants during the Mesozoic but are uncommon today.
|

|
- Angiospermae: (Total group name: Anthophyta) Cretaceous-recent) Flowering plants.
|

|
How are these related?

- Some potential synapomorphies support a Cycad - Gingko clade
- Conifers are, in any event, closer to angiosperms than to cycads
- "Gymnosperms" - non-angiosperm seed plants have traditionally been considered paraphyletic, however recent molecular analyses strongly support them as monophyletic. (Although morphology based studies don't.)
- Gnetales have traditionally been viewed as very close to the ancestry of angiosperms, but these same molecular studies nest them within gymnosperms, possibly as members of the conifers.
Stay tuned.




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}