


Key Points:
•Pterosauria represents a clade of ornithodiran archosaurs that achieved flight in the Late Triassic and survived until the very end of the Cretaceous.
•Pterosaurs were ecologically diverse; metabolically they seem to have been tachymetabolic homeothermic endotherms.
•There is no compelling evidence that they were ecologically competing with Mesozoic birds, occupying a different set of niches than birds before the K/Pg extinction.
•Diverse groups of sauropsids (reptiles) invaded the sea during the Mesozoic.
•While many Mesozoic marine sauropsids lived in near-shore environments, several were pelagic (adapted for life in the open seas). These include the superficially-dolphin-like ichthyosaurs; the diverse plesiosaurs; the marine true lizards the mosasaurs; marine crocodiliforms; marine turtles; and hesperornithine birds. (Only the hesperornithines were actually dinosaurs!)
•Many marine reptiles gave birth live directly in the water, allowing them to evolve away from any particular land-moving adaptations.
Not all reptiles of the Mesozoic were dinosaurs. There was a diversity of reptiles on land, and also in the sea and in the air.
A number of Permian & Triassic gliding reptiles, but only two powered flying groups:
Simplified cladogram of Archosauria.
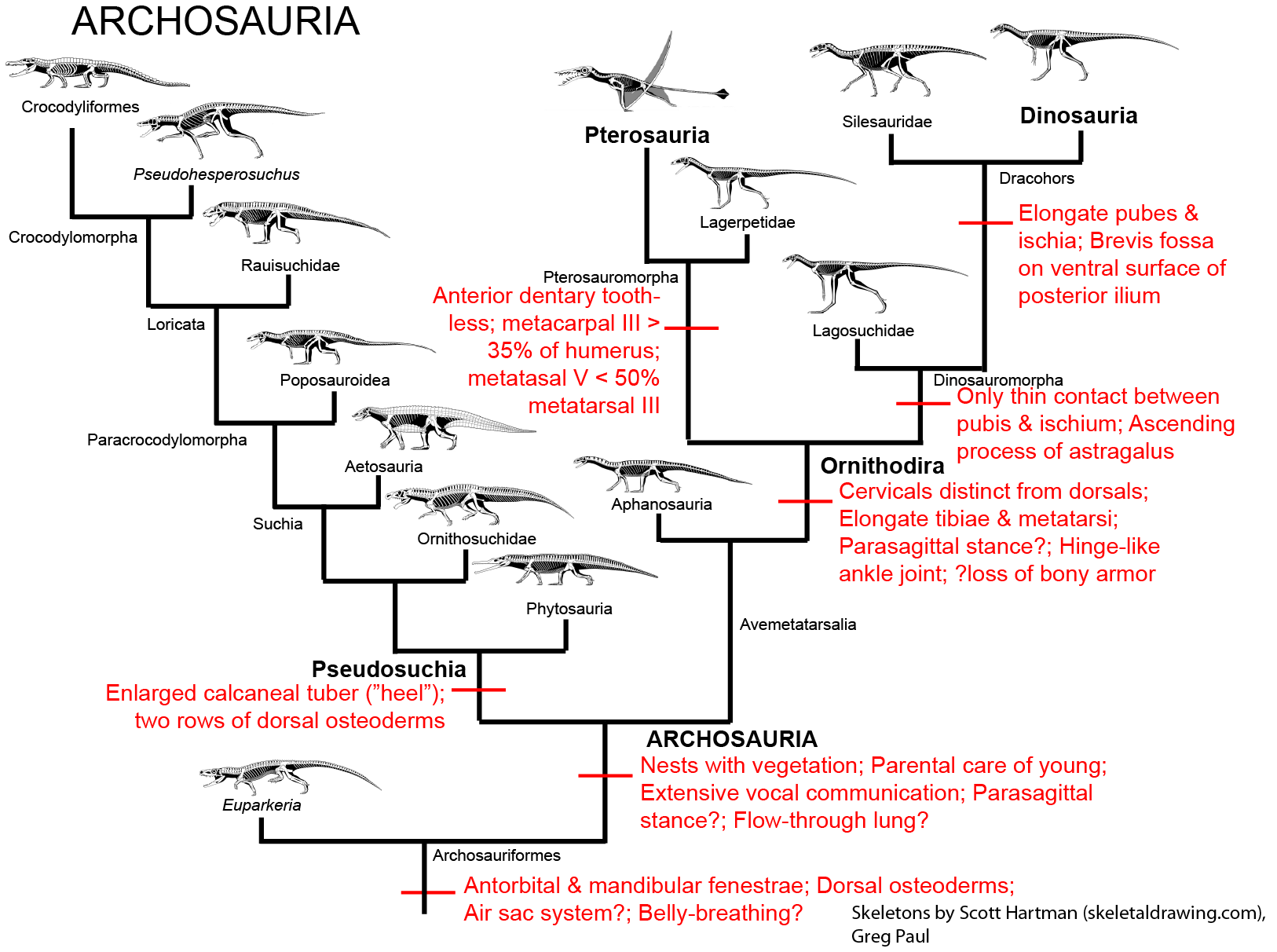
More detailed phylogeny of Avemetatarsalia, with concentration on the early dinosaurs (and using the Saurischia model).

Pterosaurs
The name Pterosauria means "wing reptile". Pterosaurs were ornithodiran archosaurs, the sister group to the dinosauromorphs. The oldest pterosaurs originated in the Late Triassic, slightly younger than the oldest known dinosaur. The last pterosaurs disappeared at the very end of the Cretaceous. Recent work shows that the lagerpetids were basal pterosauromorphs.
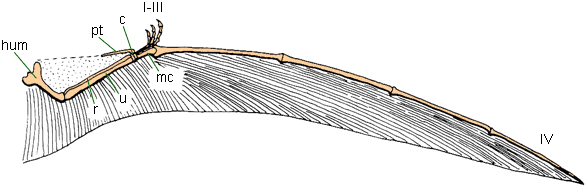
Their wing is primarily formed from digit IV of the hand: in other words, their wing finger is their ring finger! (They did not have pinkies!). But the pterosaurian wing is more complex than that. They had a specialized wrist bone (the pteroid) which controlled a flap of a "front wing". The main wing went from the tip of the finger to the ankle (at least ancestrally). There was another flap of wing running from foot to foot, underneath the tail. But the wing wasn't just skin stretched between limbs: fibers connected to muscles ran through the wing, allowing pterosaurs to reshape their wing under active control.
Pterosaur bones were very hollow: even more hollow than than the bones of birds! Like birds (and at least some other dinosaurs), the pterosaurs had complex air sacs invading vertebrae and limb bones, which lightened the skeleton as was part of an advanced respiratory system.
The muscle attachments for pterosaur wings are very well-developed, showing that they were active powered fliers and not simple gliders (as some once argued.)
The body of pterosaurs was covered with a fur named pycnofibers. This fur is not at all homologous to mammalian fur; it MIGHT be homologous with the fuzz in ornithischian and theropod dinosaurs, but uncertainty about the exact phylogenetic distribution of these features means that this is not resolved.
Basal pterosaurs are sometimes called "rhamphorhynchoids", but this is a paraphyletic grade rather than a clade. "Rhamphorhynchoids" often had long tails (at least some of which ended in diamond- or Valentine heart-shaped fins.) They do not seem to have been as effective runners on the ground as their pterodactyloid descendants, and pterosaur ptrace fossils are almost unknown until Pterodactyloidea appears.
There are many varieties of "rhamphorhynchoids": early ones have diverse tooth shapes and a skull reminiscent of theropod dinosaurs; others have short frog-shaped faces; still others have long grasping teeth possible for snatching fish. "Rhamphorhynchoids" range from about the size of a robin to that of an eagle. The last of the "rhamphorhynchoid" pterosaurs disappear in the later Early Cretaceous.
Darwinopterus and other wukongopterids of the Middle and Late Jurassic represent a transition to the true Pterodactyloidea ("wing fingers"). Pterodactyloids first appear near the beginning of the Late Jurassic. They differ from basal pterosaurs by a reduced tail size, a much longer palm of the hand (metacarpus), and a split in the hindwing (so it no longer runs from foot-to-foot). The latter two changes (as well as some in the hips) points to them being better movers on the ground, and indeed pterodactyloid ptracks are fairly common in the Late Jurassic and Cretaceous. Pterodactyloids (at least: maybe other pterosaurs!) launched by using their very powerful forelimbs to push them into the air. (In nearly all other fliers and gliders, the initial launch is from the hindlimbs.)
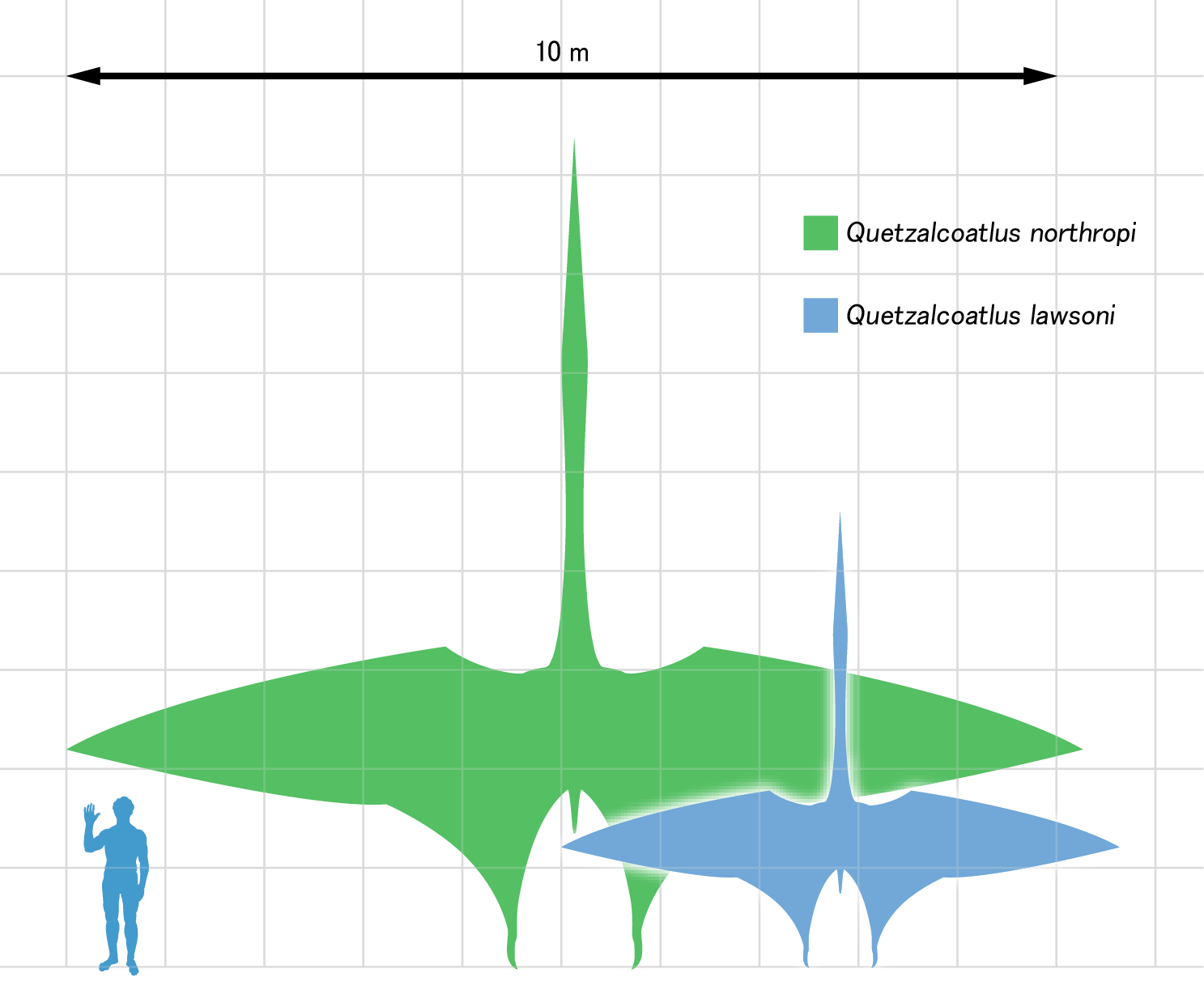
Many pterodactyloids (and some rhamphorhynchoids, too) had crests on their head: some were bony, others made of firm or soft tissue. There appears in at least some cases to be sexual dimorphism in the size and shape of the crests. Pterodactyloid size ranges were far greater than "rhamphorhynchoids": the smallest were sparrow-sized but the largest were by far the largest fliers of all time: 12-14 m wingspans and the height of a giraffe!
Although some once argued that pterosaurs walked bipedally, but both anatomical and trace-fossil studies demonstrate that they walked quadupedally (almost gorilla-like). Trackway evidence for pterosaurs are rare in the Triassic and Early and Middle Jurassic, but become common in the Late Jurassic onward. This coincides with the rise of pterodactyloids, which show more adaptations for ground-walking.
Pterosaur diets varied: some may have been insectivores, others piscivores (fish-eaters), still others filter feeders (like modern flamingos), some herbivores, some durophagous (eating hard objects), and a few carnivorous. None seem to have been true raptorial forms in the sense of hawks and eagles, though.
Pterosaur eggs were definitely more leathery than at least some dinosaurs' (which were crispy, as in the living dinosaur [aka bird] eggs). The embryos show the same wing proportions as free-living babies, and it seems that they could fly from birth. However, this does not mean that they abandoned their young: after all, crocodilians, birds, and mammals with freely-moving young often have extensive parental care.
Like non-avian dinosaurs, pterosaurs grew faster than living cold-blooded animals, achieving full body size in less than a decade. As noted above, they seem to have had complex air sacs that could pump air through their body efficiently. Their growth rates, respiration, fur, ability to use powered flight, bone texture, and other features point to them having a fully warm-blooded metabolism. (In fact, it had been proposed they were warm-blooded even before Dinosauria was discovered!)
Throughout Earth History, many reptiles (and other amniotes) have returned to the sea:
All marine amniotes share similar sets of problems:
First reptiles to return to an aquatic life were mesosaurs:
Most primitive relatives of Mesozoic marine sauropsids were similar in general form (long needle-like teeth, webbed hands and feet, deep tail, some terrestrial ability, probably shore-dwelling or fresh-water) to mesosaurs, but later forms become more specialized for life in the sea.
Many different clades of Mesozoic marine sauropsids, from almost every clade:
We will cover the most diverse and highly specialized forms: euryapsids (esp. ichthyosaurs and plesiosaurs), mosasaurs, and marine turtles.
A reminder of amniote phylogeny:
:

Euryapsida: A clade of diapsid reptile in which the infratemporal fenestra is closed, so that only the supratemporal fenestra is open. The position of euryapsids is uncertain: some have found them to be lepidosauromorphs, some as archosauromorphs, some as closer to turtles. The position shown in the cladogram above has them outside the Lepidosauromorpha/Archosauromorpha clade Sauria, which has been found in a number of recent studies. (It is worth noting that some analyses don't find a monophyletic Euryapsida, and instead find that ichthyosauromorphs (ichthyosaurs and hupeheuchians) plus thallatosaurs form one clade, separate from Sauropterygia (placodonts, pachypleurosaurs, nothosaurs, and pistosaurs (including plesiosaurs)).
Euryapsids:
Ichthyopterygia (ichthyosaurs):
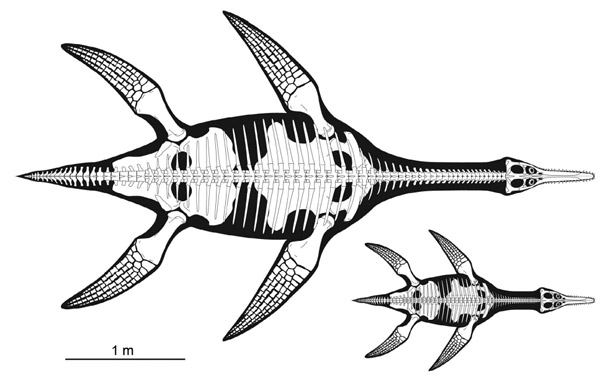
Plesiosauria (plesiosaurs):
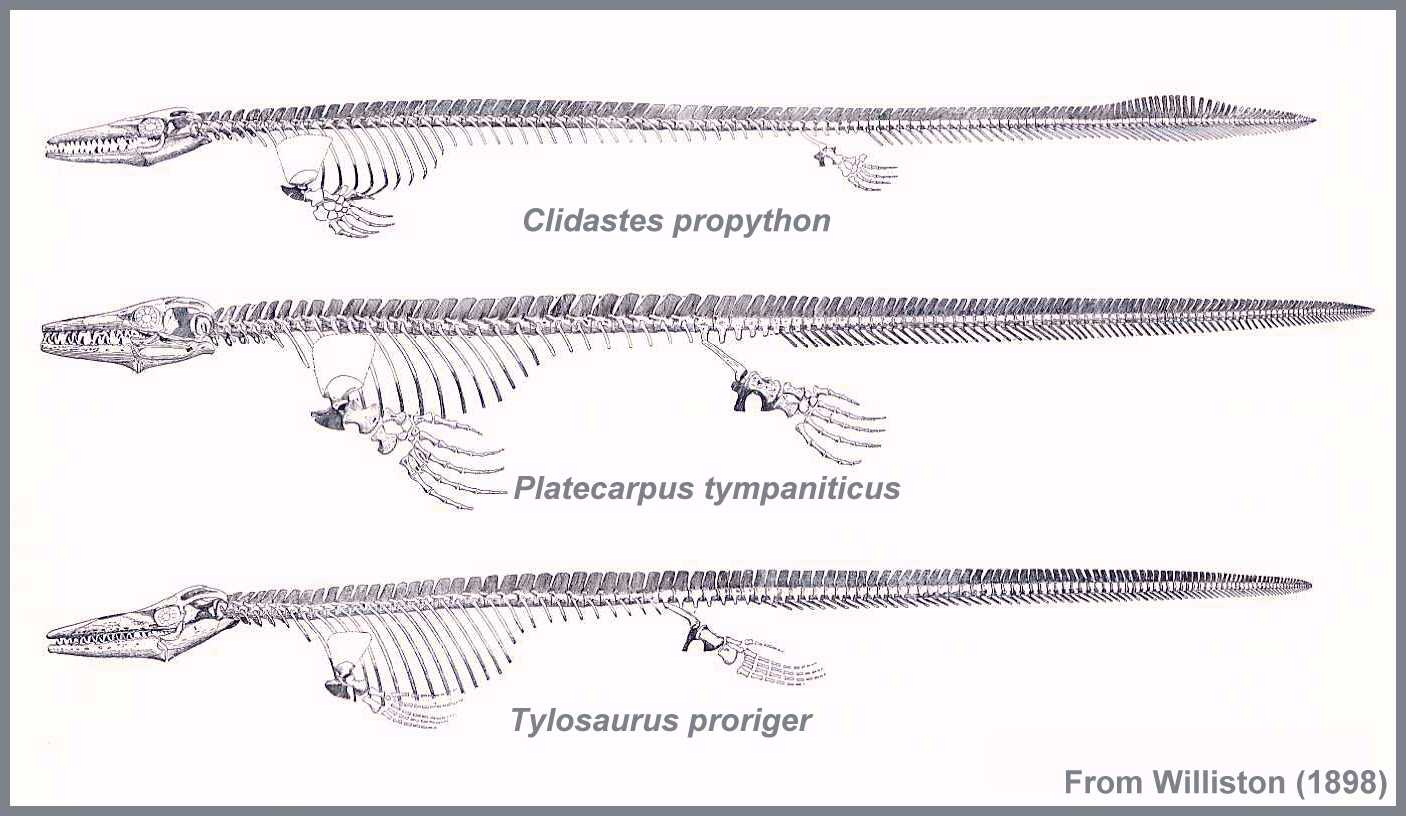
Mosasauroidea (mosasaurs):
Marine turtles (Chelonioidea and Angolchelonia):
The Hot-Blooded Ichthyosaurs?: Recent work on isotopic data and the bone growth rate data strongly suggest that the pistosaurs (plesiosaurs and the closest relatives) and derived (Jurassic and Cretaceous) ichthyosaurs were fully endothermic, that mosasaurids may have been as well, and that thallatosuchian crocs had a higher metabolic rate than typical endotherms. One trait shared by these but not found in other marine reptiles (for whom the data do NOT support endothermy) is that they were all large fast-swimming pelagic hunters. Interestingly, several groups of living fish (tuna; billfish like marlins, swordfish, and sailfish; and some lamniform sharks such as makos and great whites) are fast-swimming large pelagic hungers and also endothermic. It appears that in the marine realm the transition to endothermy was not through miniaturization of animals with intermediate metabolic rates, but rather the evolution of a fast fish-chasing lifestyle among larger animals.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}